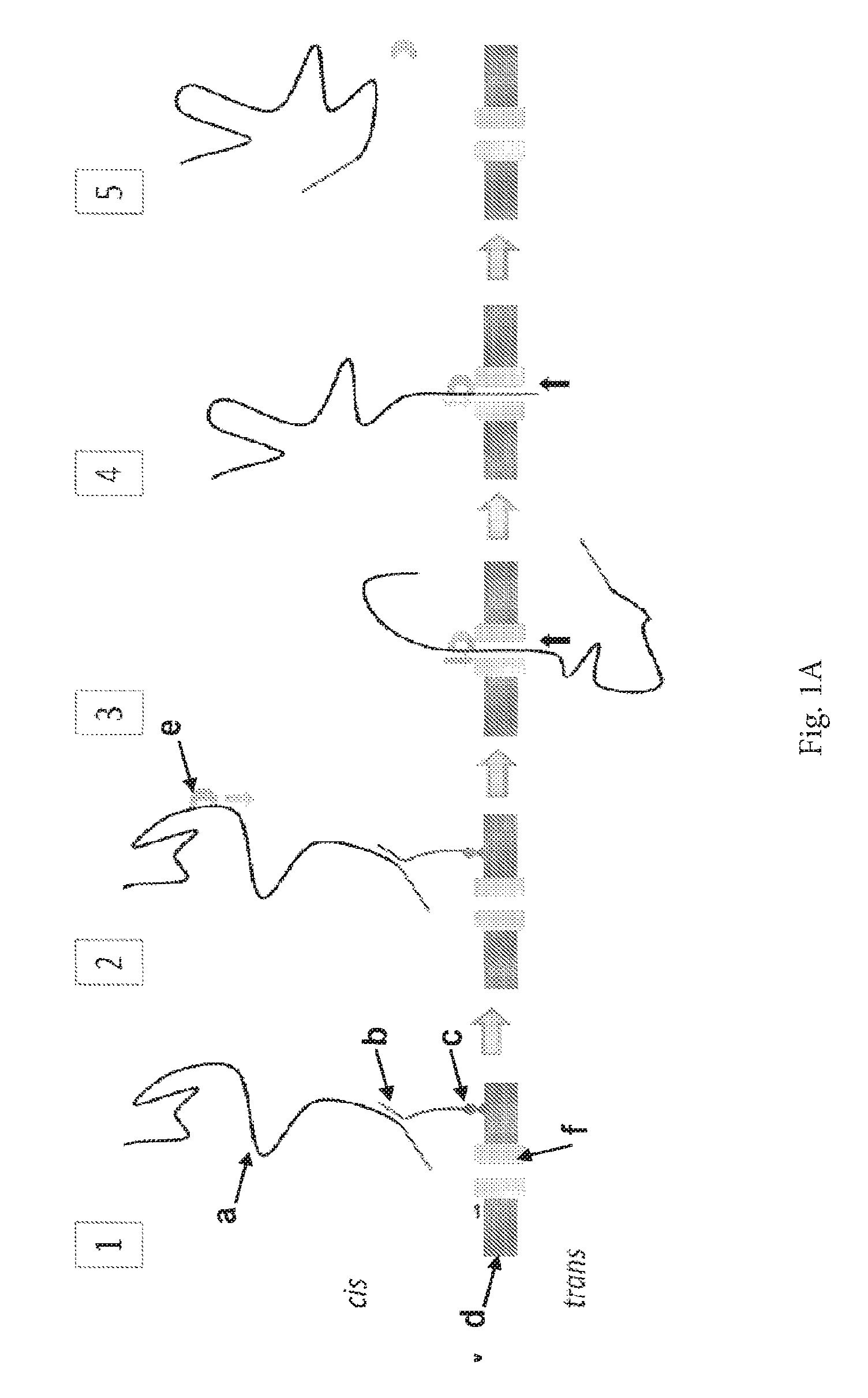
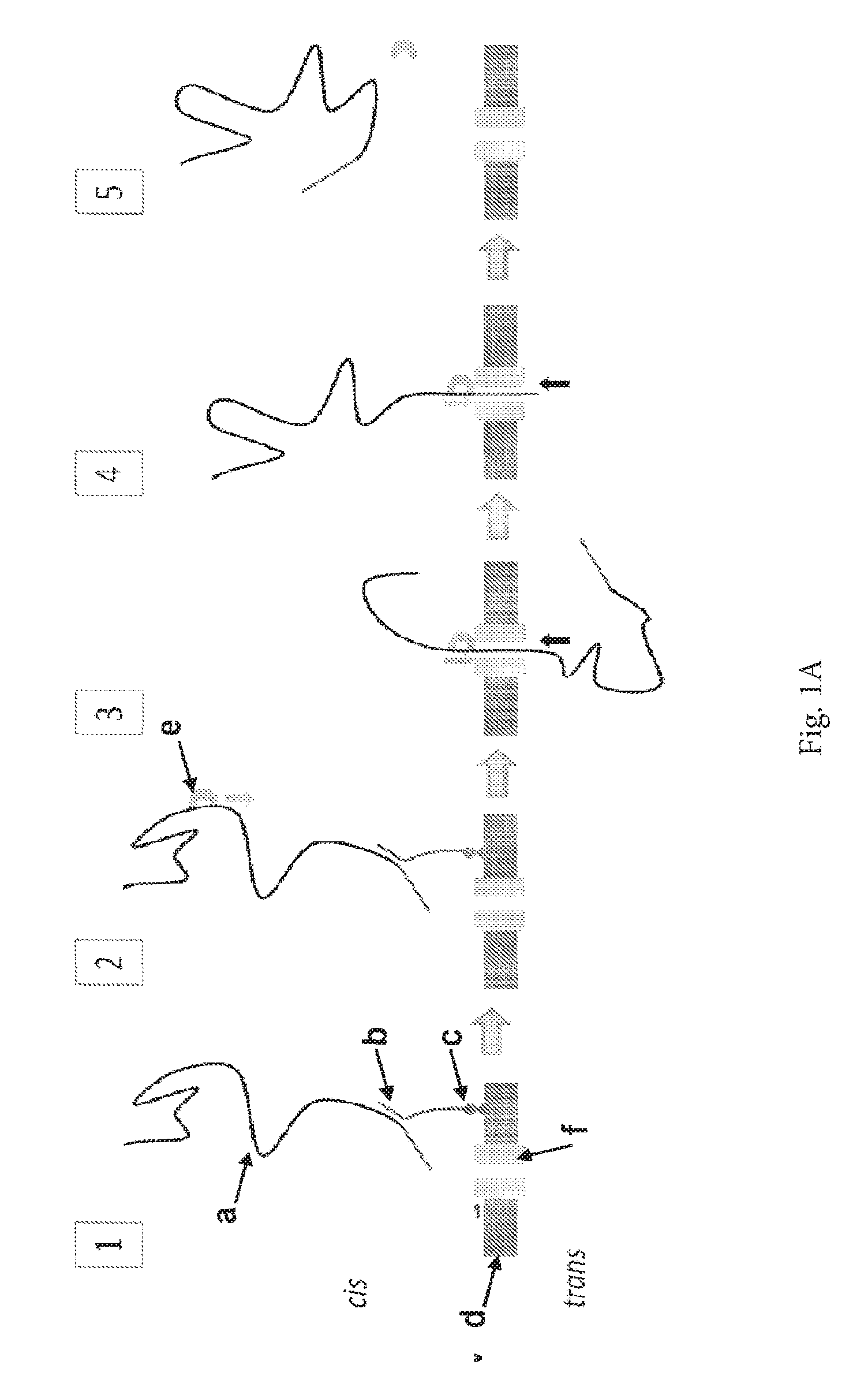
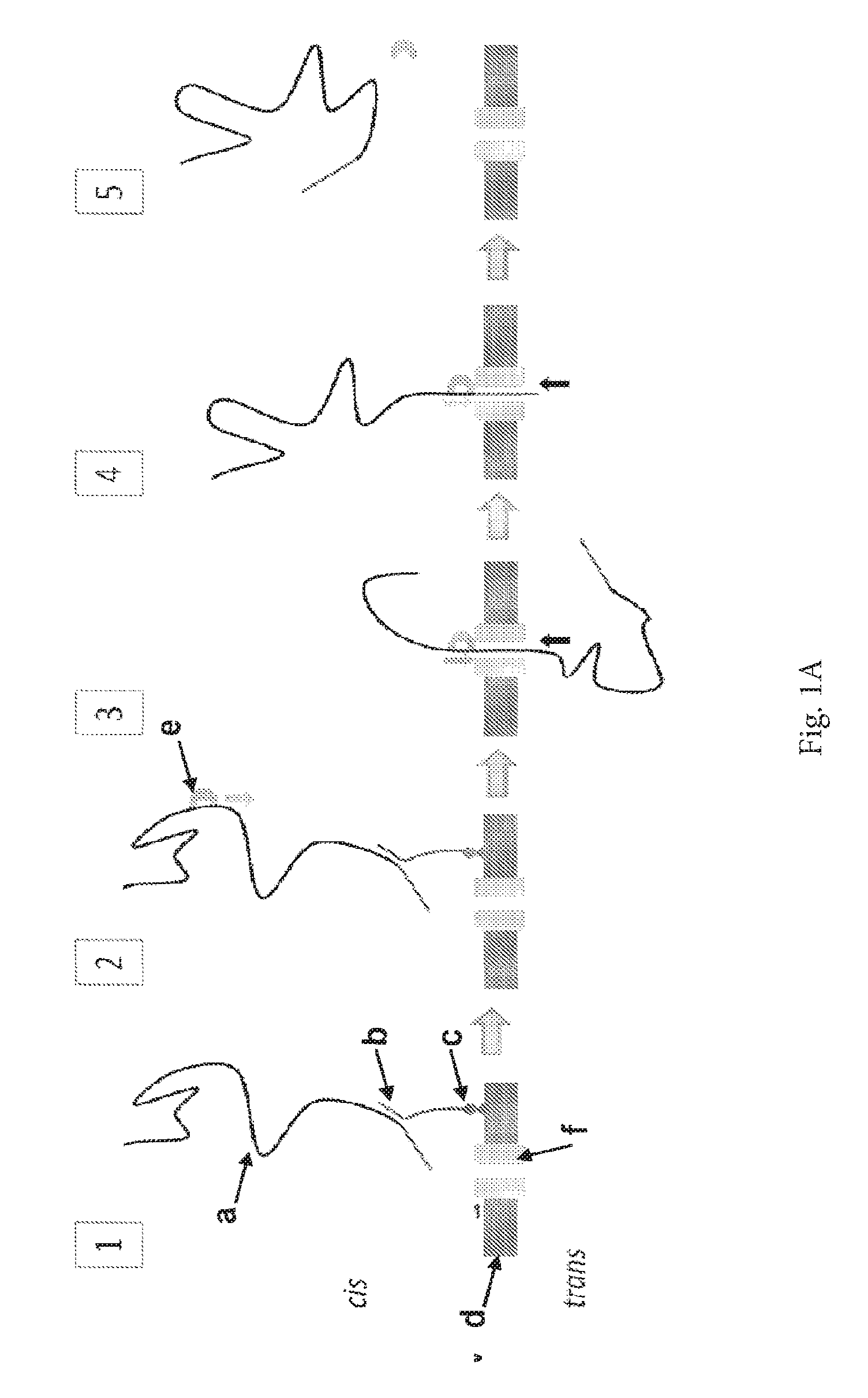
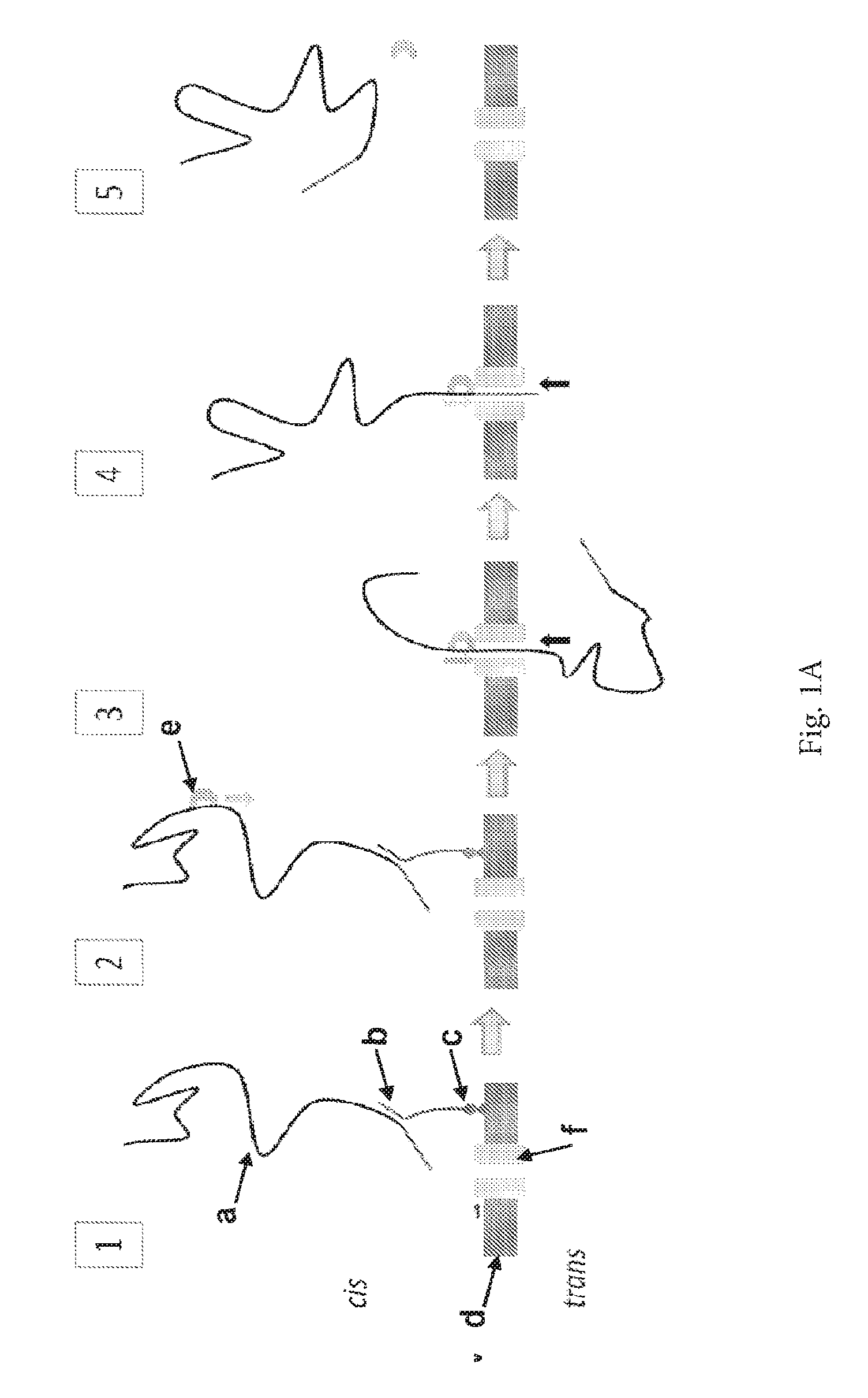
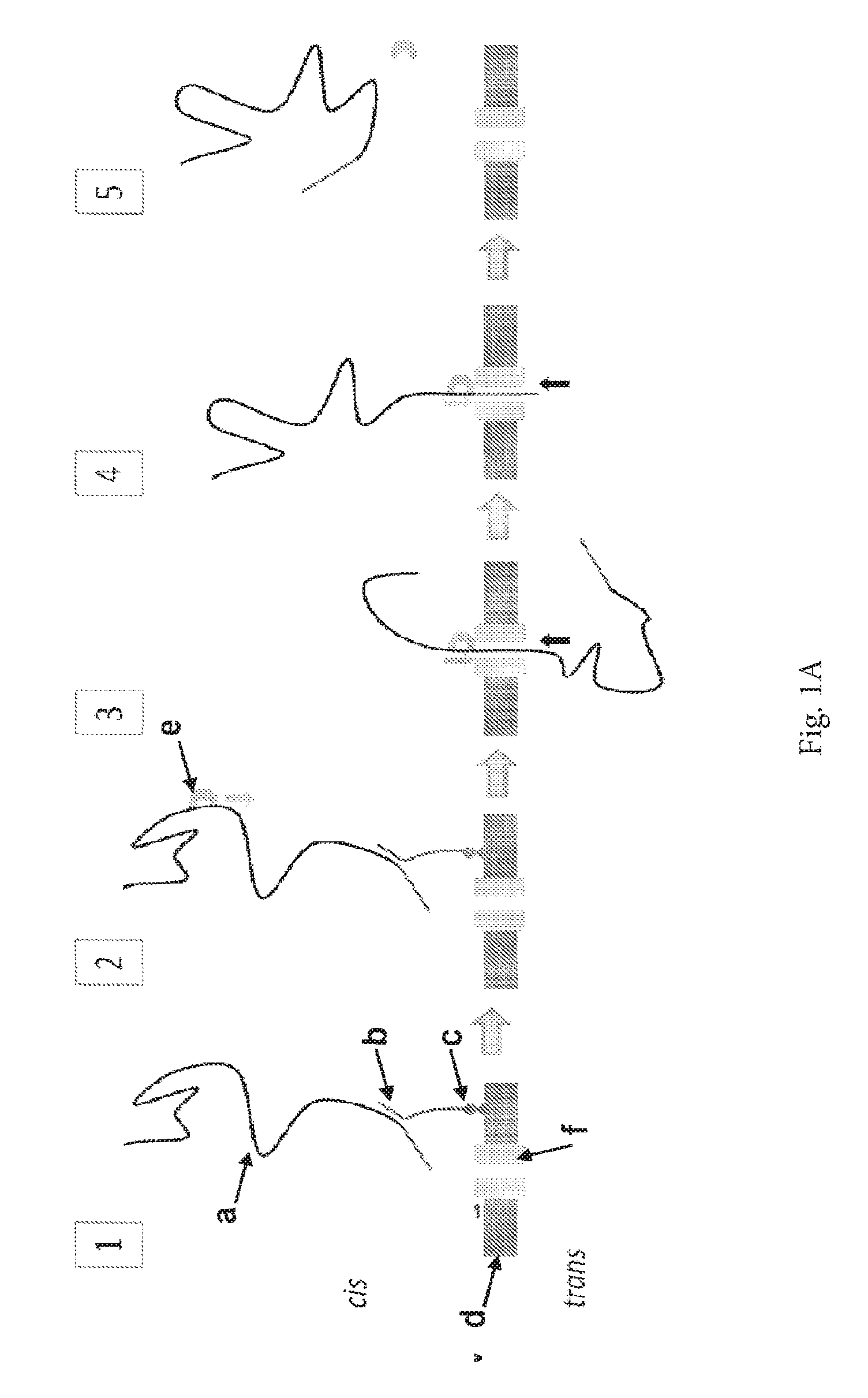
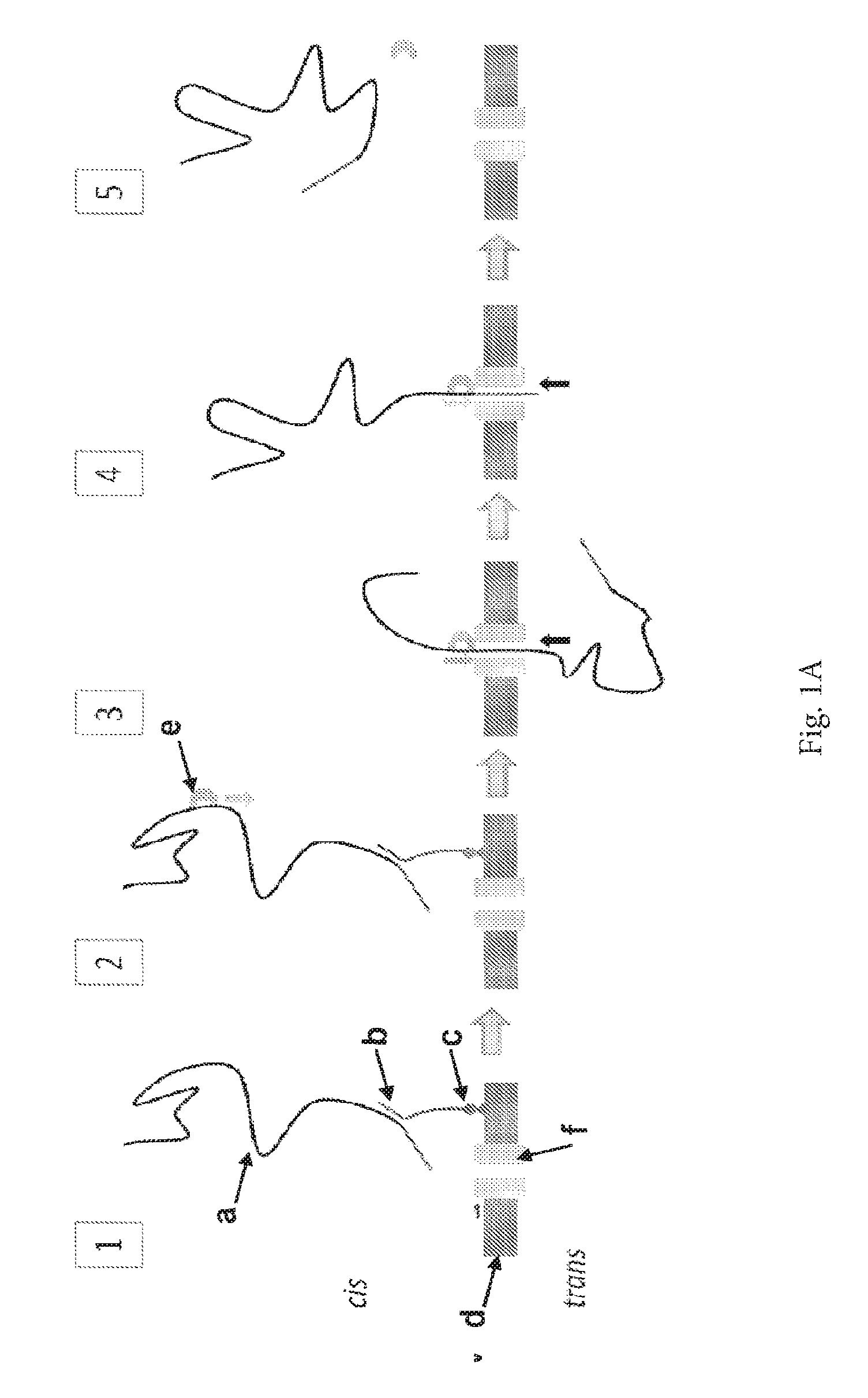
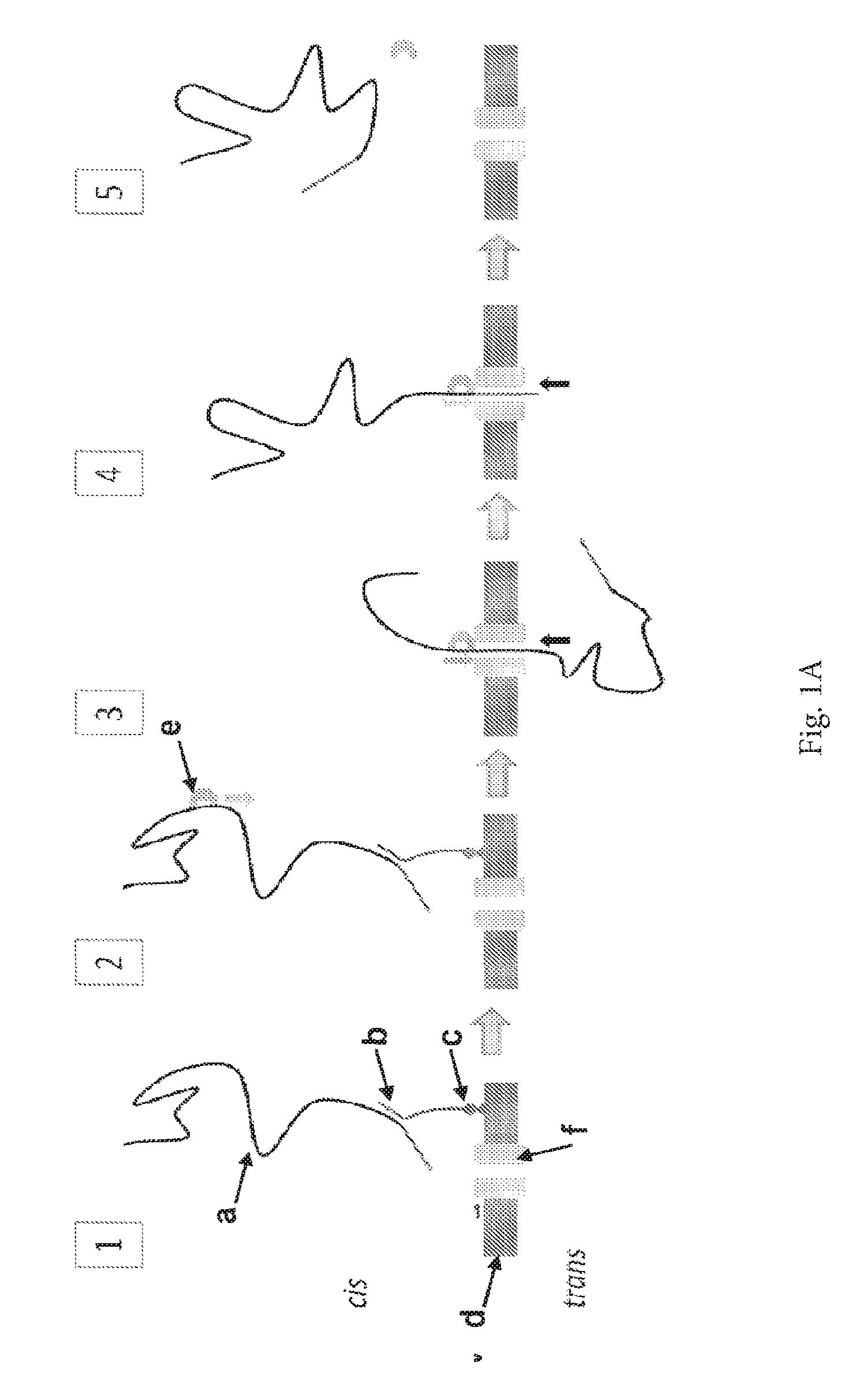
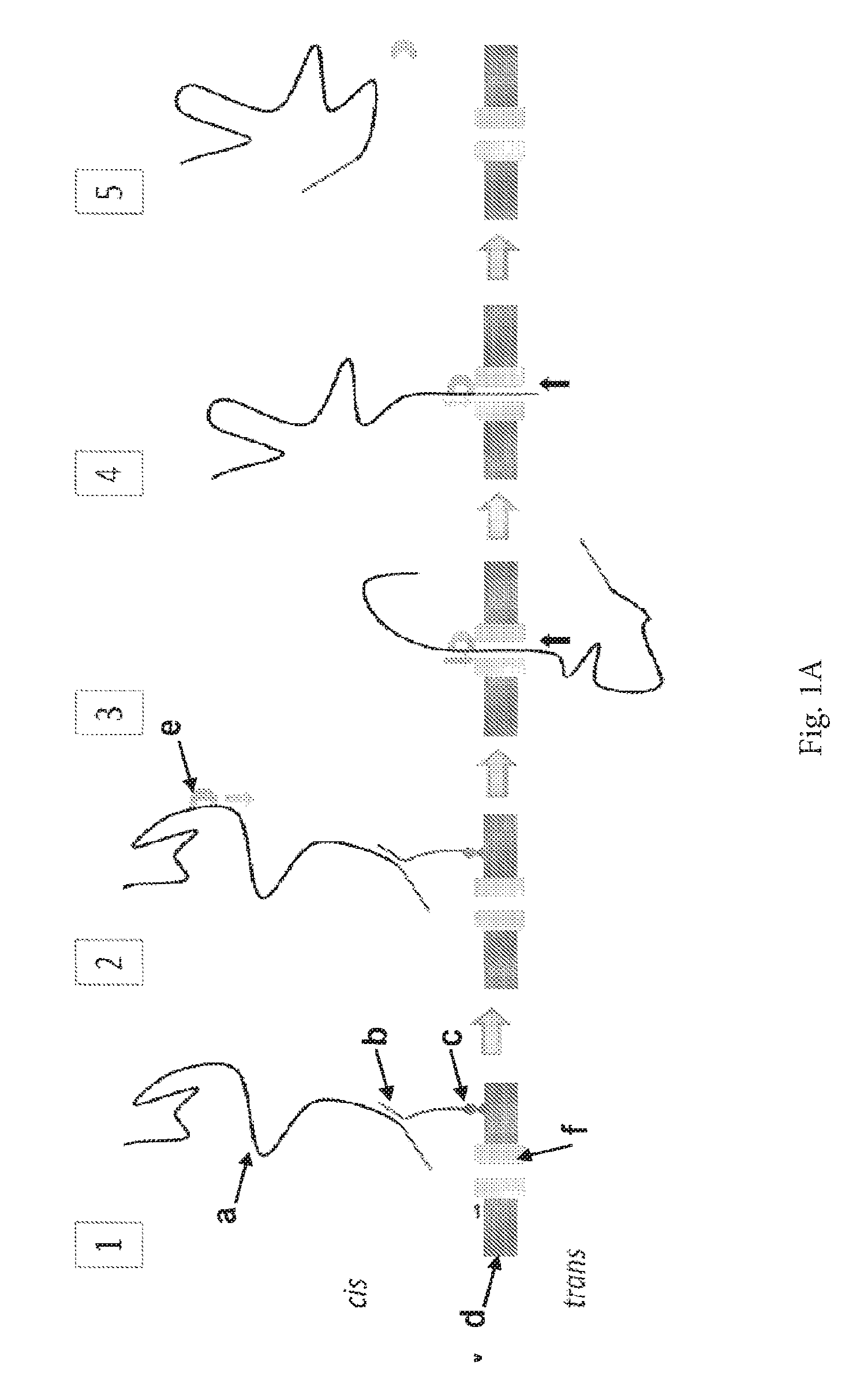
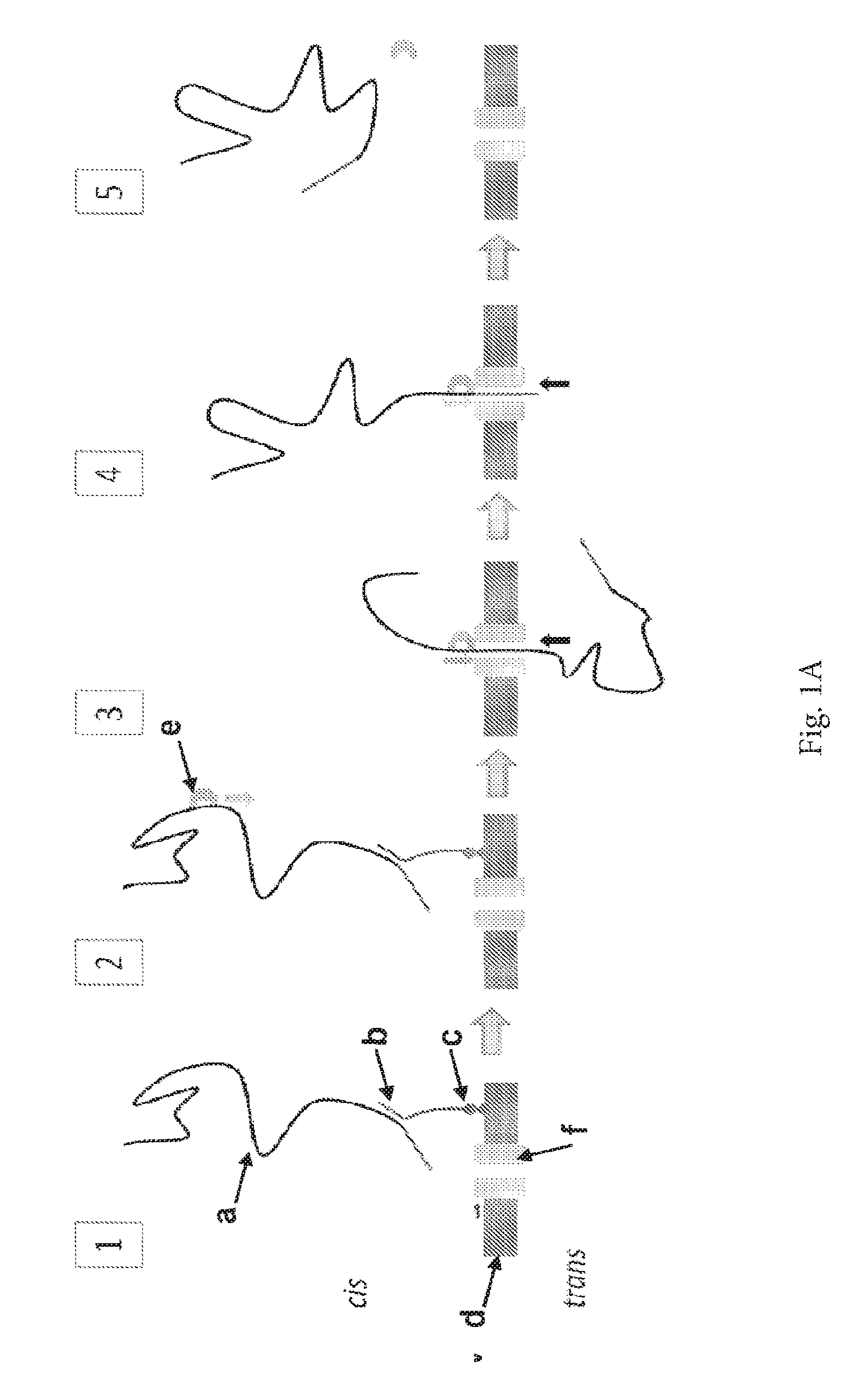
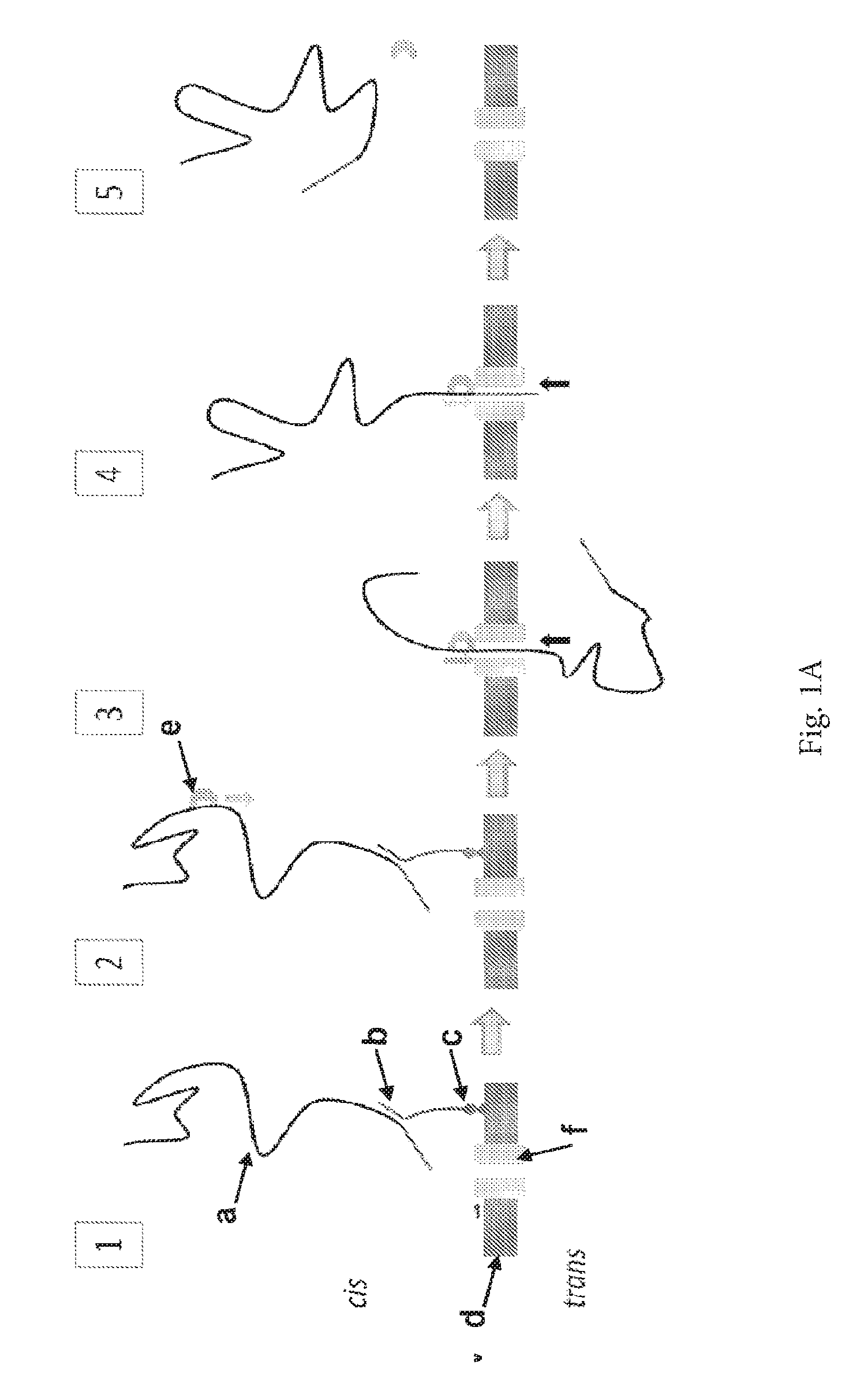
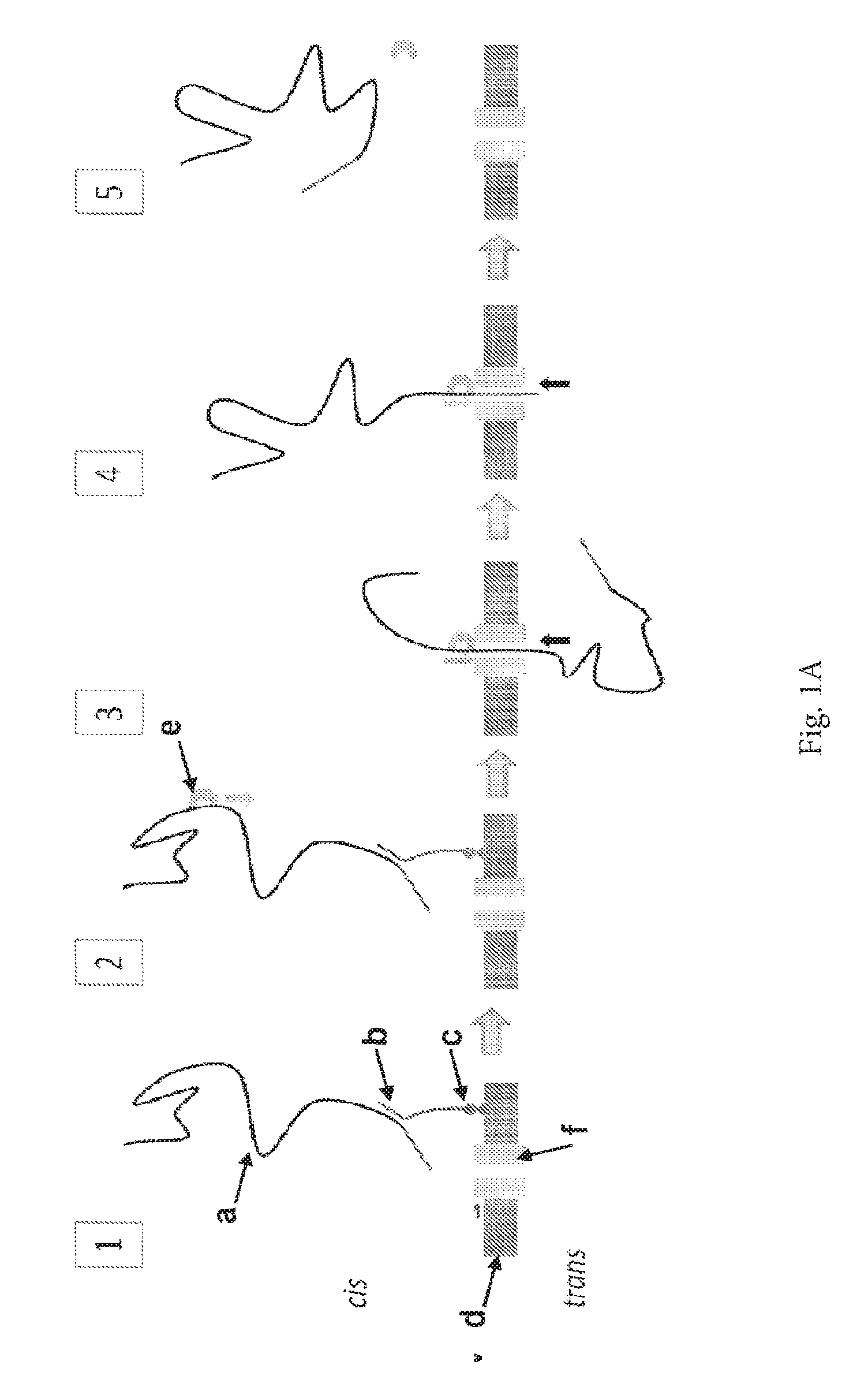
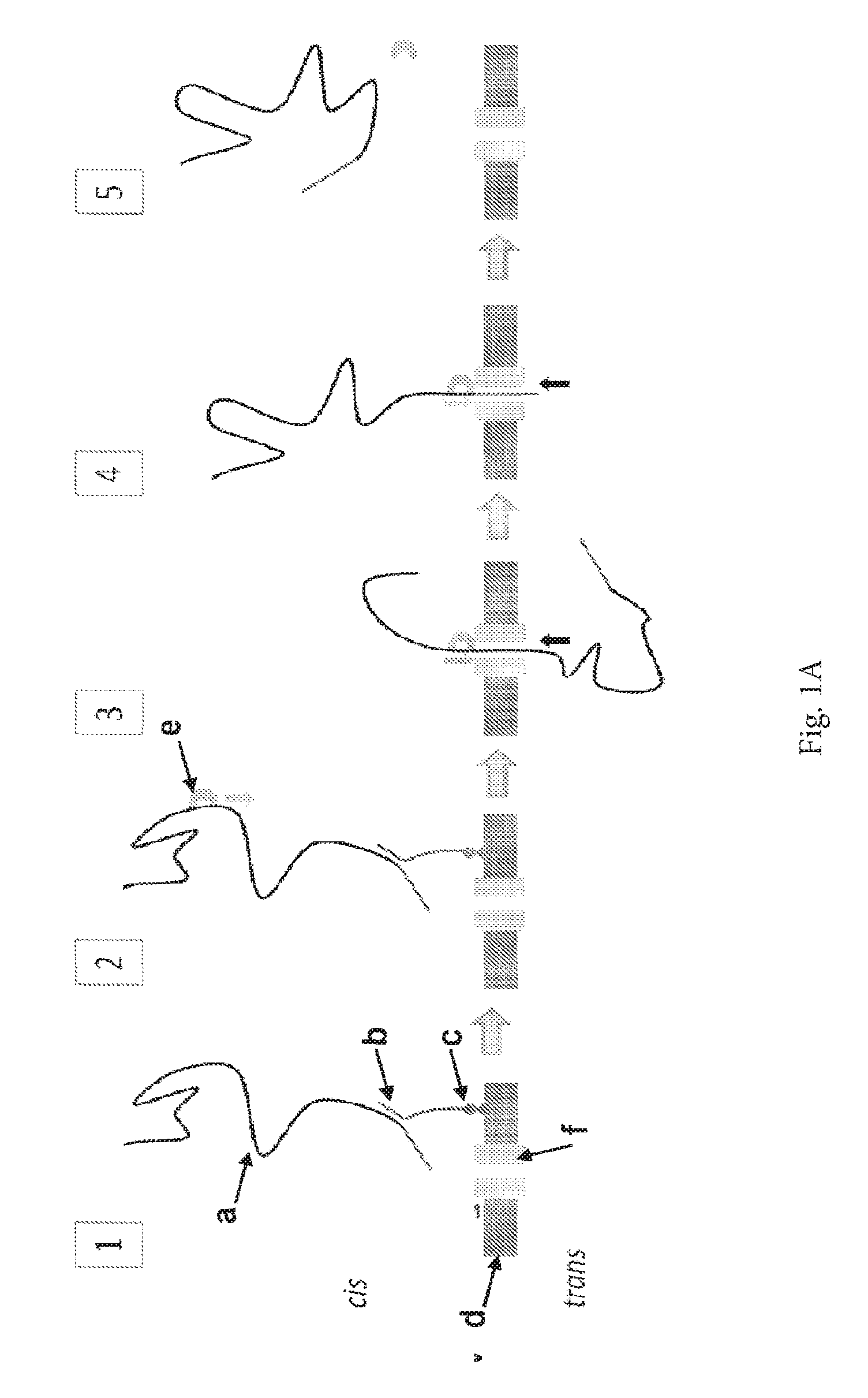
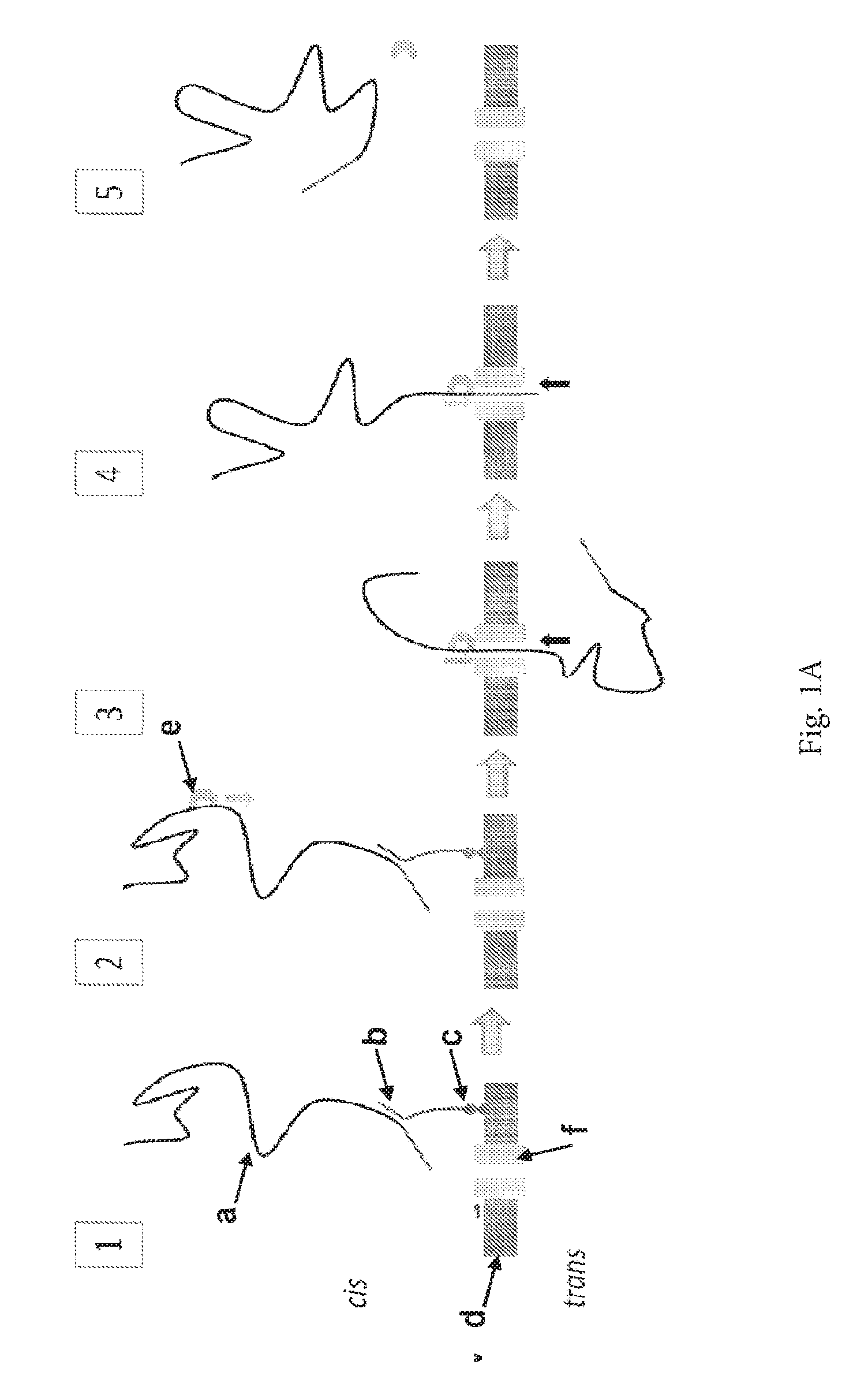
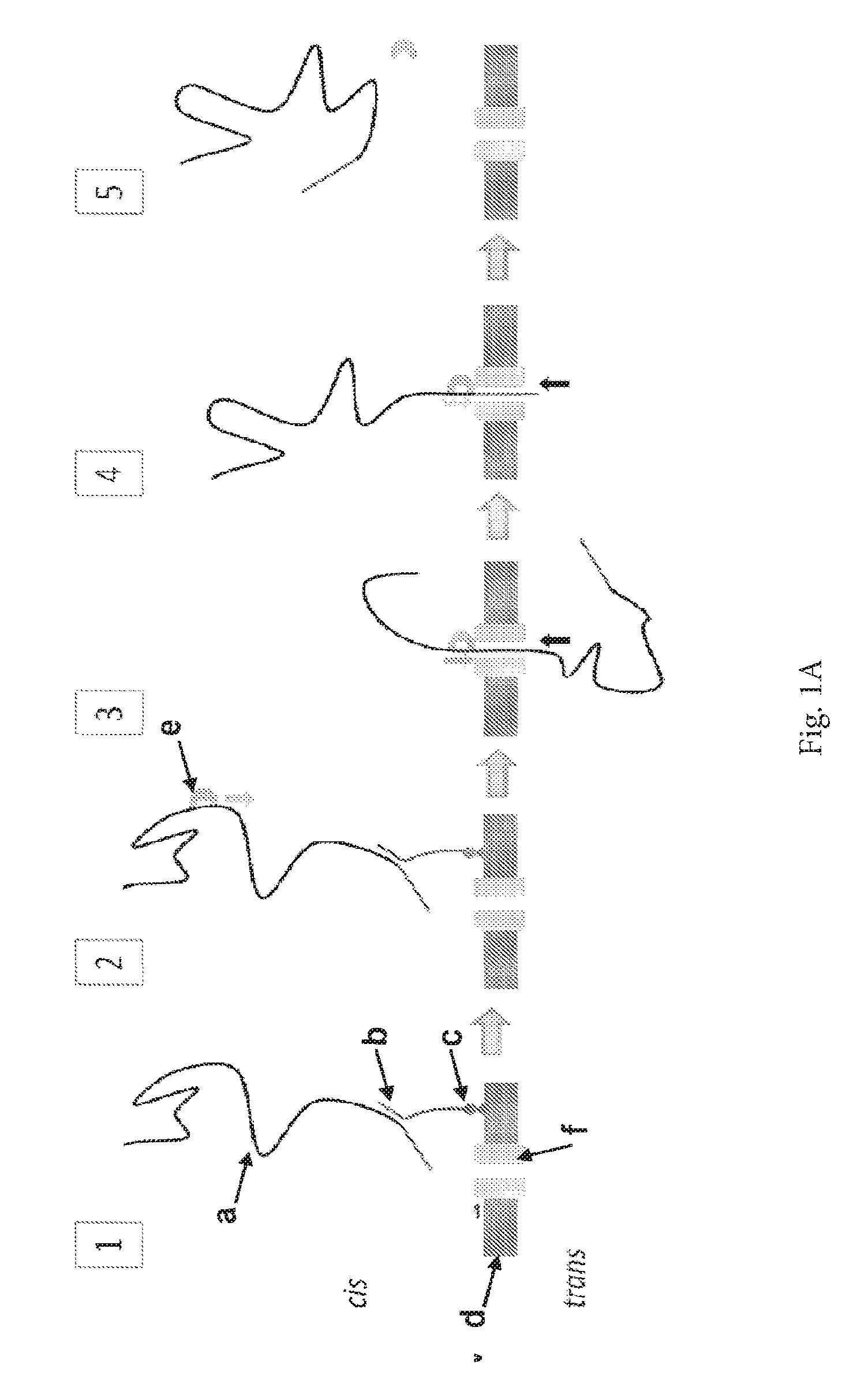
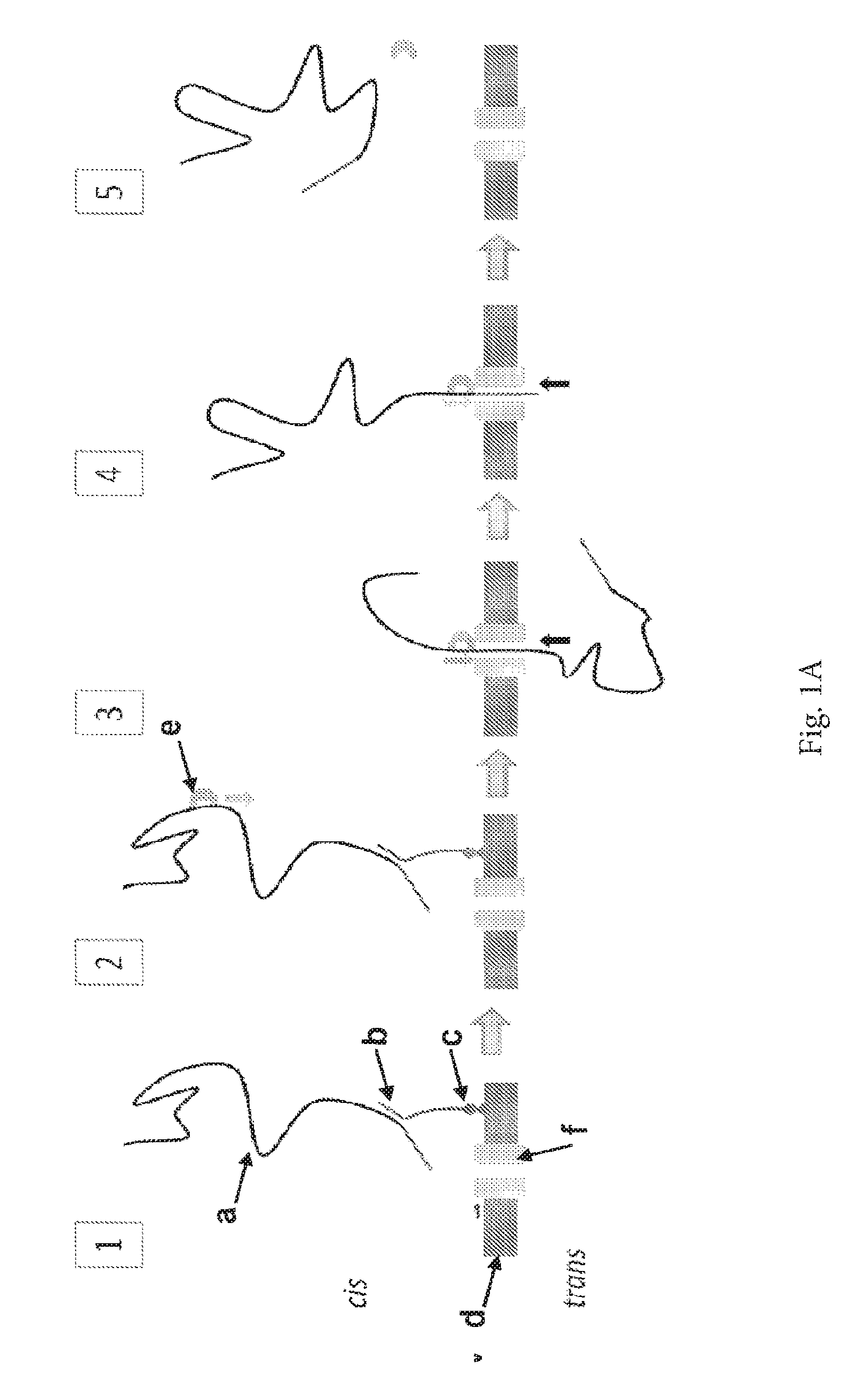
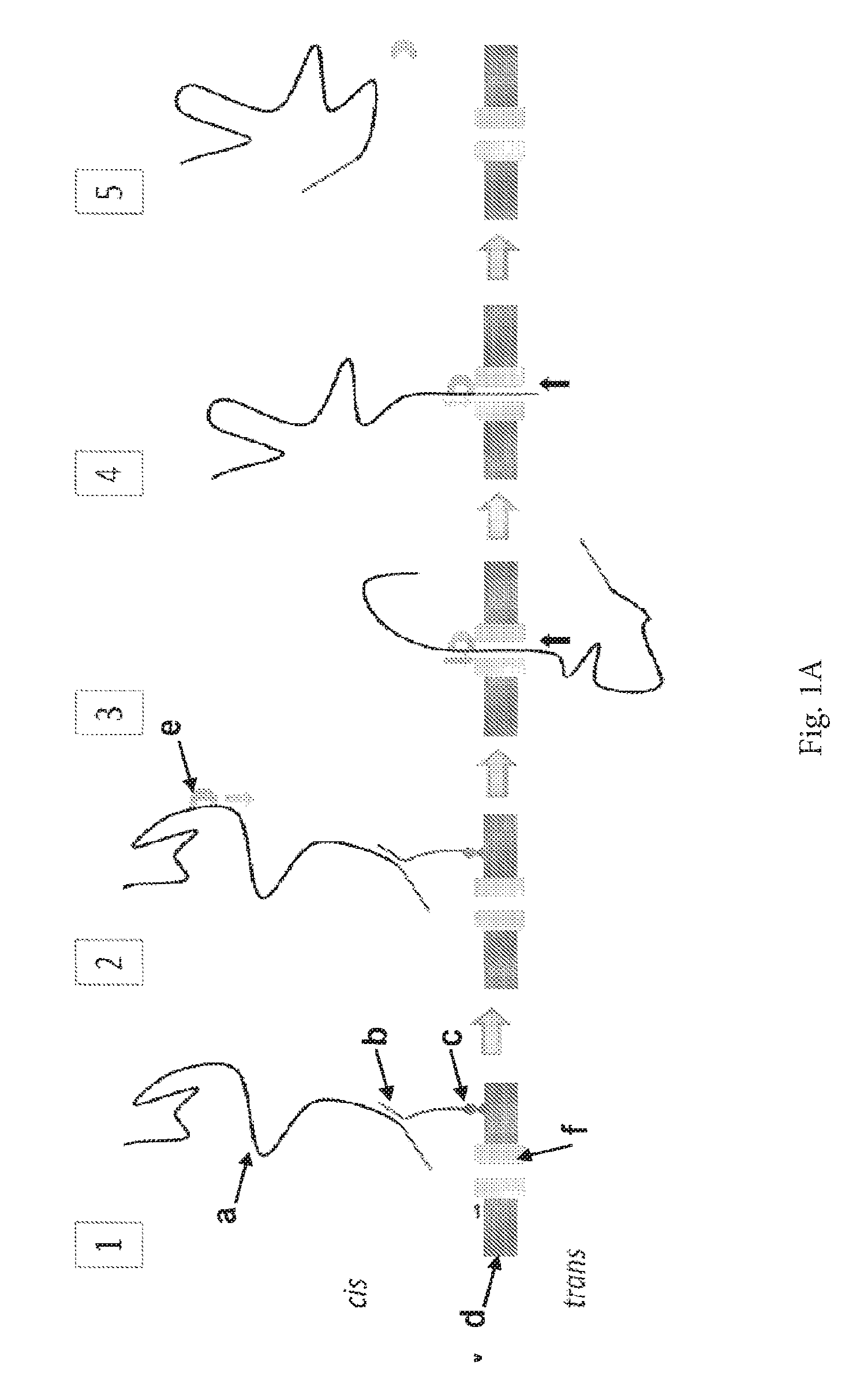
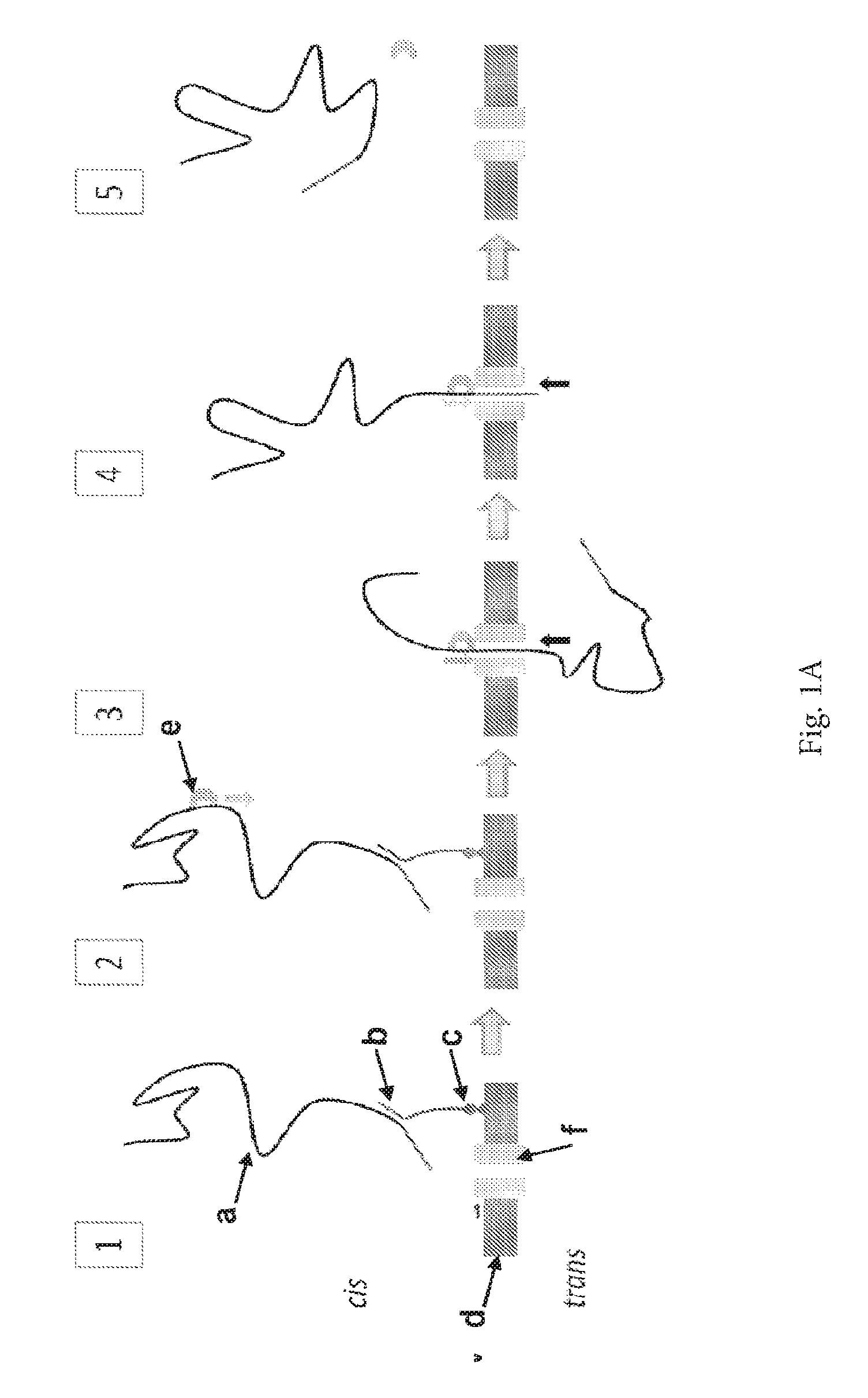
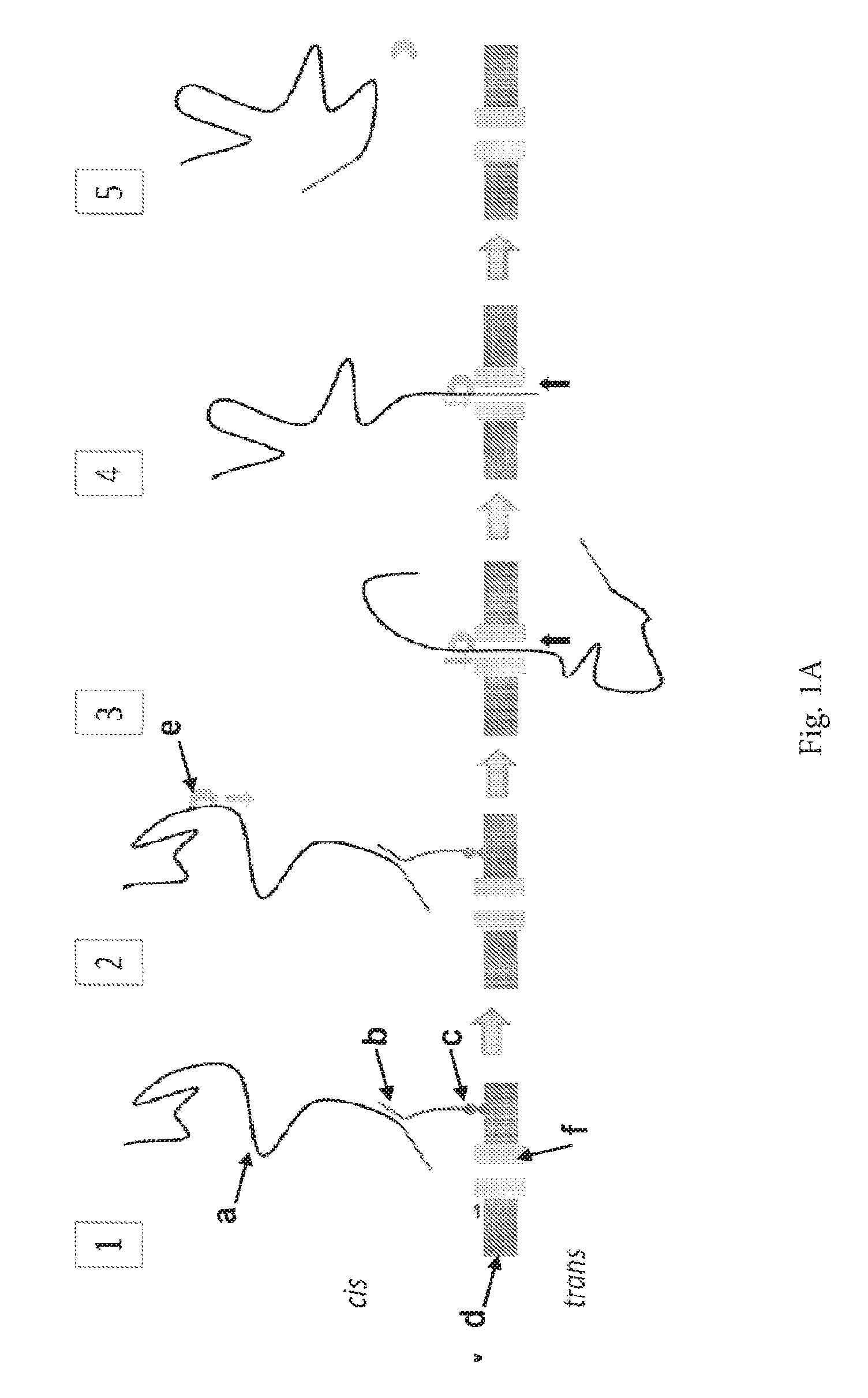
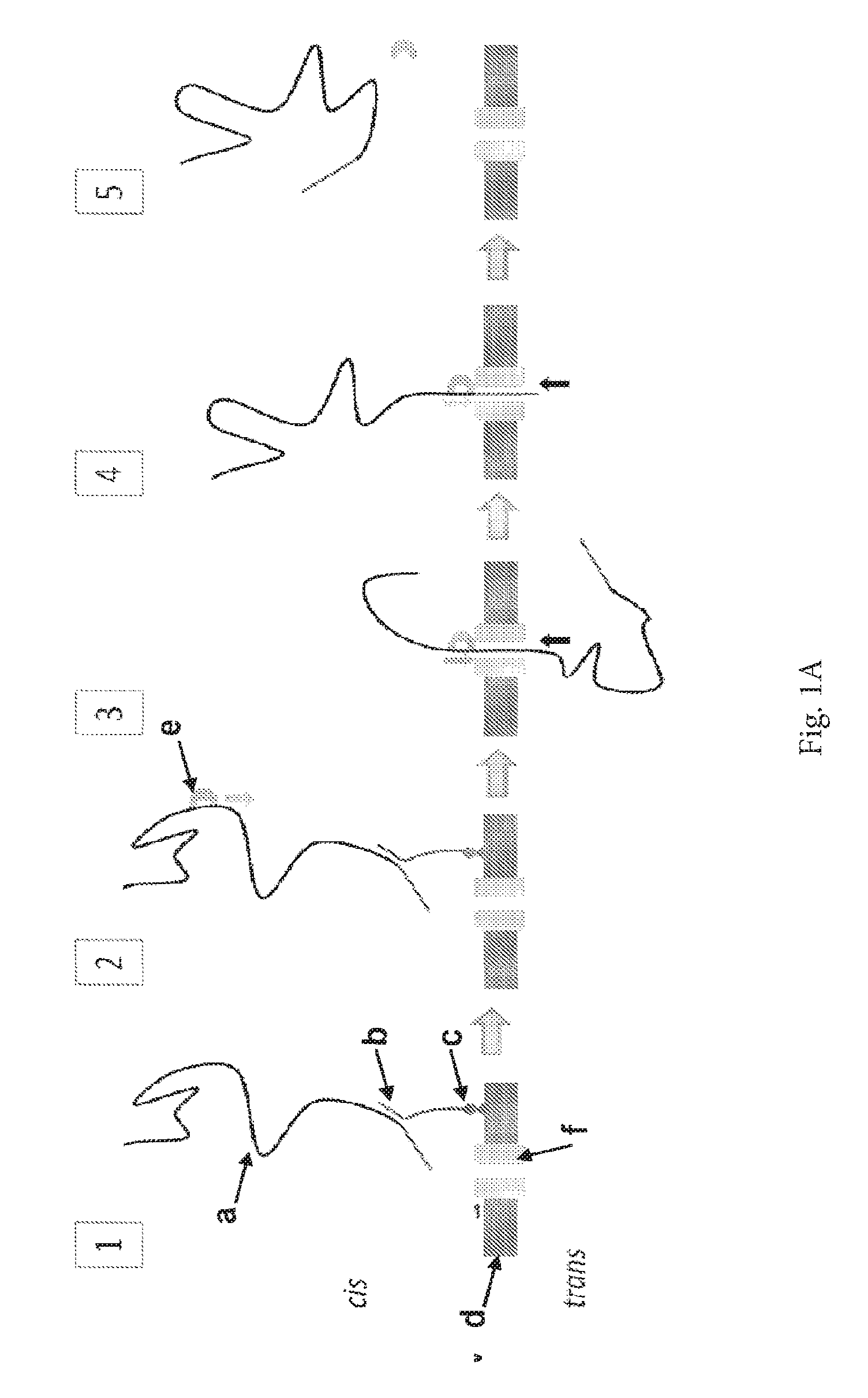
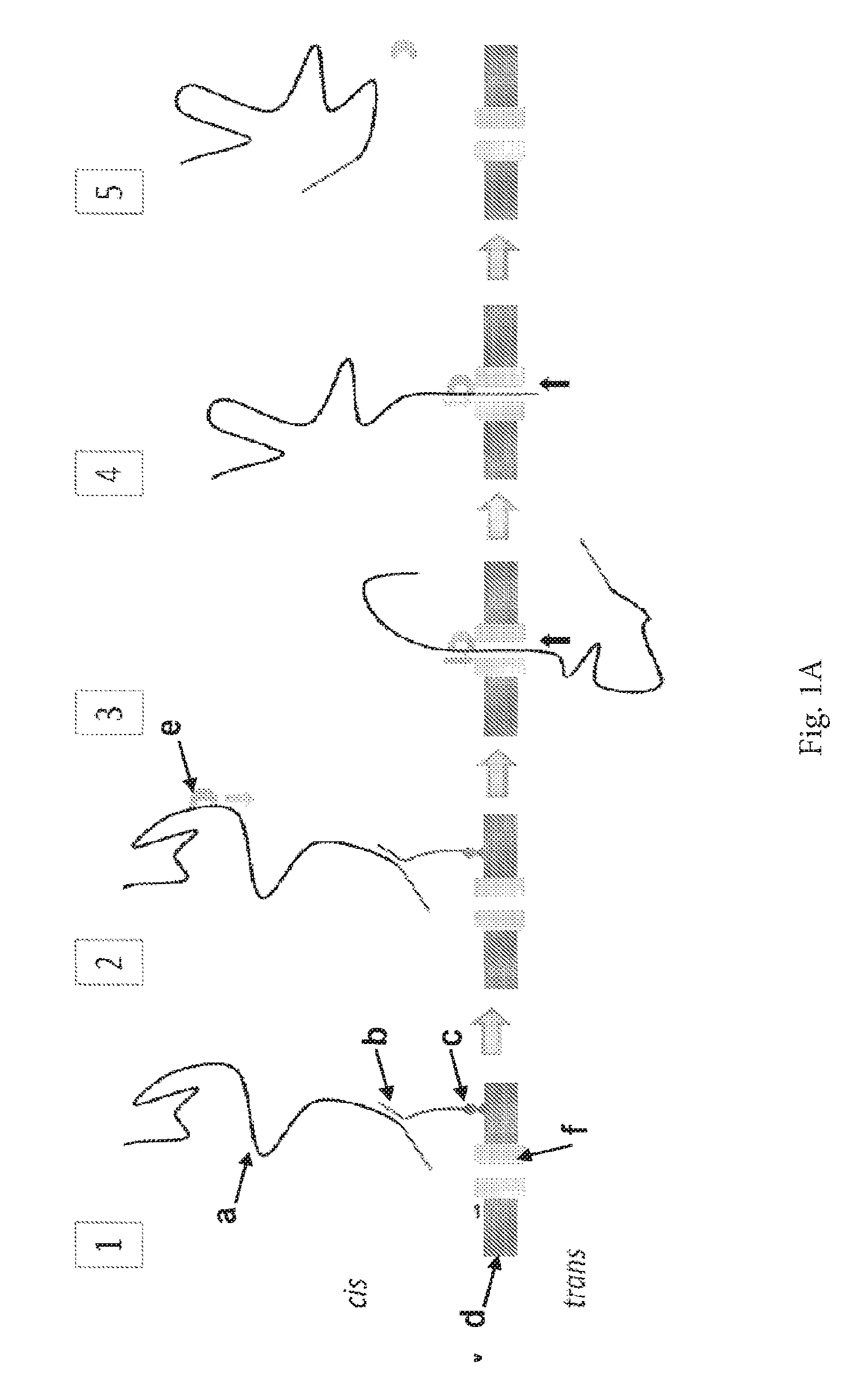
ENZYME METHOD
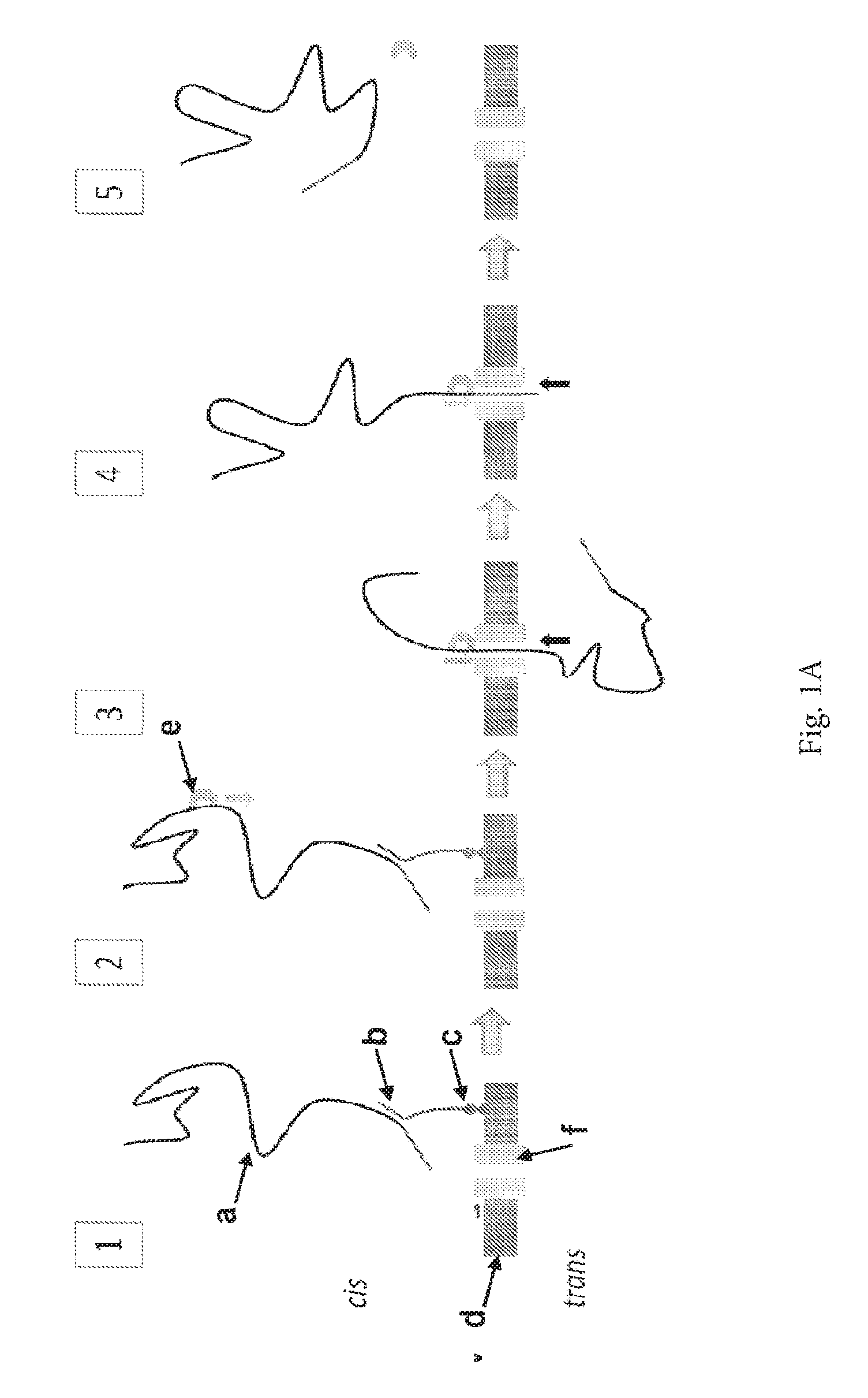
The invention relates to a new method of characterising a target polynucleotide. The method uses a pore and a Hel308 helicase or a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide. The helicase or molecular motor controls the movement of the target polynucleotide through the pore. There is currently a need for rapid and cheap polynucleotide (e.g. DNA or RNA) sequencing and identification technologies across a wide range of applications. Existing technologies are slow and expensive mainly because they rely on amplification techniques to produce large volumes of polynucleotide and require a high quantity of specialist fluorescent chemicals for signal detection. Transmembrane pores (nanopores) have great potential as direct, electrical biosensors for polymers and a variety of small molecules. In particular, recent focus has been given to nanopores as a potential DNA sequencing technology. When a potential is applied across a nanopore, there is a change in the current flow when an analyte, such as a nucleotide, resides transiently in the barrel for a certain period of time. Nanopore detection of the nucleotide gives a current change of known signature and duration. In the “Strand Sequencing” method, a single polynucleotide strand is passed through the pore and the identity of the nucleotides are derived. Strand Sequencing can involve the use of a nucleotide handling protein to control the movement of the polynucleotide through the pore. The inventors have demonstrated that a Hel308 helicase can control the movement of a polynucleotide through a pore especially when a potential, such as a voltage, is applied. The helicase is capable of moving a target polynucleotide in a controlled and stepwise fashion against or with the field resulting from the applied voltage. Surprisingly, the helicase is capable of functioning at a high salt concentration which is advantageous for characterising the polynucleotide and, in particular, for determining its sequence using Strand Sequencing. This is discussed in more detail below. Accordingly, the invention provides a method of characterising a target polynucleotide, comprising: (a) contacting the target polynucleotide with a transmembrane pore and a Hel308 helicase such that the helicase controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore; and (b) measuring one or more characteristics of the target polynucleotide during one or more interactions and thereby characterising the target polynucleotide. The invention also provides:
The inventors have also demonstrated that a molecular motor which is capable of binding to a target polynucleotide at an internal nucleotide can control the movement of the polynucleotide through a pore especially when a potential, such as a voltage, is applied. The motor is capable of moving the target polynucleotide in a controlled and stepwise fashion against or with the field resulting from the applied voltage. Surprisingly, when the motor is used in the method of the invention it is possible to control the movement of an entire strand of target polynucleotide through a nanopore. This is advantageous for characterising the polynucleotide and, in particular, for determining its sequence using Strand Sequencing. Hence, the invention also provides a method of characterising a target polynucleotide, comprising: (a) contacting the target polynucleotide with a transmembrane pore and a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide such that the molecular motor controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore; and (b) measuring one or more characteristics of the target polynucleotide during one or more interactions and thereby characterising the target polynucleotide. SEQ ID NO: 1 shows the codon optimised polynucleotide sequence encoding the MS-B1 mutant MspA monomer. This mutant lacks the signal sequence and includes the following mutations: D90N, D91N, D93N, D118R, D134R and E139K. SEQ ID NO: 2 shows the amino acid sequence of the mature form of the MS-B1 mutant of the MspA monomer. This mutant lacks the signal sequence and includes the following mutations: D90N, D91N, D93N, D118R, D134R and E139K. SEQ ID NO: 3 shows the polynucleotide sequence encoding one subunit of α-hemolysin-E111N/K147N (α-HL-NN; Stoddart et al., PNAS, 2009; 106(19): 7702-7707). SEQ ID NO: 4 shows the amino acid sequence of one subunit of α-HL-NN. SEQ ID NOs: 5 to 7 shows the amino acid sequences of MspB, C and D. SEQ ID NO: 8 shows the amino acid sequence of the Hel308 motif. SEQ ID NO: 9 shows the amino acid sequence of the extended Hel308 motif. SEQ ID NOs: 10 to 58 show the amino acid sequences of the Hel308 helicases and motifs in Table 5. SEQ ID NOs: 59 to 74 show the sequences used in the Examples. SEQ ID NO: 75 shows the sequence of Hel308 Dth in the alignmenton on page 57 onwards. SEQ ID NO: 76 shows the sequence of Hel308 Mmar in the alignment on page 57 onwards. SEQ ID NO: 77 shows the sequence of Hel308 Nth in the alignment on page 57 onwards. SEQ ID NO: 78 shows the consensus sequence in the alignment on page 57 onwards. It is to be understood that different applications of the disclosed products and methods may be tailored to the specific needs in the art. It is also to be understood that the terminology used herein is for the purpose of describing particular embodiments of the invention only, and is not intended to be limiting. In addition as used in this specification and the appended claims, the singular forms “a”, “an”, and “the” include plural referents unless the content clearly dictates otherwise. Thus, for example, reference to “a pore” includes two or more such pores, reference to “a helicase” includes two or more such helicases, reference to “a polynucleotide” includes two or more such polynucleotides, and the like. All publications, patents and patent applications cited herein, whether supra or infra, are hereby incorporated by reference in their entirety. The invention provides a method of characterising a target polynucleotide. The method comprises contacting the target polynucleotide with a transmembrane pore and a Hel308 helicase such that the helicase controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore. One or more characteristics of the target polynucleotide are then measured using standard methods known in the art. Steps (a) and (b) are preferably carried out with a potential applied across the pore. As discussed in more detail below, the applied potential typically results in the formation of a complex between the pore and the helicase. The applied potential may be a voltage potential. Alternatively, the applied potential may be a chemical potential. An example of this is using a salt gradient across the lipid membrane. A salt gradient is disclosed in Holden et al., J Am Chem Soc. 2007 Jul. 11; 129(27):8650-5. In some instances, the current passing through the pore during one or more interactions is used to determine the sequence of the target polynucleotide. This is Strand Sequencing. The method has several advantages. First, the inventors have surprisingly shown that Hel308 helicases have a surprisingly high salt tolerance and so the method of the invention may be carried out at high salt concentrations. In the context of Strand Sequencing, a charge carrier, such as a salt, is necessary to create a conductive solution for applying a voltage offset to capture and translocate the target polynucleotide and to measure the resulting sequence-dependent current changes as the polynucleotide passes through the pore. Since the measurement signal is dependent on the concentration of the salt, it is advantageous to use high salt concentrations to increase the magnitude of the acquired signal. High salt concentrations provide a high signal to noise ratio and allow for currents indicative of the presence of a nucleotide to be identified against the background of normal current fluctuations. For Strand Sequencing, salt concentrations in excess of 100 mM are ideal and salt concentrations of 1 M and above are preferred. The inventors have surprisingly shown that Hel308 helicases will function effectively at salt concentrations as high as, for example, 2 M. Second, when a voltage is applied, Hel308 helicases can surprisingly move the target polynucleotide in two directions, namely with or against the field resulting from the applied voltage. Hence, the method of the invention may be carried out in one of two preferred modes. Different signals are obtained depending on the direction the target polynucleotide moves through the pore, ie in the direction of or against the field. This is discussed in more detail below. Third, Hel308 helicases typically move the target polynucleotide through the pore one nucleotide at a time. Hel308 helicases can therefore function like a single-base ratchet. This is of course advantageous when sequencing a target polynucleotide because substantially all, if not all, of the nucleotides in the target polynucleotide may be identified using the pore. Fourth, Hel308 helicases are capable of controlling the movement of single stranded polynucleotides and double stranded polynucleotides. This means that a variety of different target polynucleotides can be characterised in accordance with the invention. Fifth, Hel308 helicases appear very resistant to the field resulting from applied voltages. The inventors have seen very little movement of the polynucleotide under an “unzipping” condition. This is important because it means that there are no complications from unwanted “backwards” movements when moving polynucleotides against the field resulting from an applied voltage. Sixth, Hel308 helicases are easy to produce and easy to handle. Their use therefore contributed to a straightforward and less expensive method of sequencing. The method of the invention is for characterising a target polynucleotide. A polynucleotide, such as a nucleic acid, is a macromolecule comprising two or more nucleotides. The polynucleotide or nucleic acid may comprise any combination of any nucleotides. The nucleotides can be naturally occurring or artificial. One or more nucleotides in the target polynucleotide can be oxidized or methylated. One or more nucleotides in the target polynucleotide may be damaged. One or more nucleotides in the target polynucleotide may be modified, for instance with a label or a tag. The target polynucleotide may comprise one or more spacers. A nucleotide typically contains a nucleobase, a sugar and at least one phosphate group. The nucleobase is typically heterocyclic. Nucleobases include, but are not limited to, purines and pyrimidines and more specifically adenine, guanine, thymine, uracil and cytosine. The sugar is typically a pentose sugar. Nucleotide sugars include, but are not limited to, ribose and deoxyribose. The nucleotide is typically a ribonucleotide or deoxyribonucleotide. The nucleotide typically contains a monophosphate, diphosphate or triphosphate. Phosphates may be attached on the 5′ or 3′ side of a nucleotide. Nucleotides include, but are not limited to, adenosine monophosphate (AMP), guanosine monophosphate (GMP), thymidine monophosphate (TMP), uridine monophosphate (UMP), cytidine monophosphate (CMP), cyclic adenosine monophosphate (cAMP), cyclic guanosine monophosphate (cGMP), deoxyadenosine monophosphate (dAMP), deoxyguanosine monophosphate (dGMP), deoxythymidine monophosphate (dTMP), deoxyuridine monophosphate (dUMP) and deoxycytidine monophosphate (dCMP). The nucleotides are preferably selected from AMP, TMP, GMP, CMP, UMP, dAMP, dTMP, dGMP or dCMP. A nucleotide may be abasic (i.e. lack a nucleobase). The polynucleotide may be single stranded or double stranded. At least a portion of the polynucleotide is preferably double stranded. The polynucleotide can be a nucleic acid, such as deoxyribonucleic acid (DNA) or ribonucleic acid (RNA). The target polynucleotide can comprise one strand of RNA hybridized to one strand of DNA. The polynucleotide may be any synthetic nucleic acid known in the art, such as peptide nucleic acid (PNA), glycerol nucleic acid (GNA), threose nucleic acid (TNA), locked nucleic acid (LNA) or other synthetic polymers with nucleotide side chains. The whole or only part of the target polynucleotide may be characterised using this method. The target polynucleotide can be any length. For example, the polynucleotide can be at least 10, at least 50, at least 100, at least 150, at least 200, at least 250, at least 300, at least 400 or at least 500 nucleotide pairs in length. The polynucleotide can be 1000 or more nucleotide pairs, 5000 or more nucleotide pairs in length or 100000 or more nucleotide pairs in length. The target polynucleotide is present in any suitable sample. The invention is typically carried out on a sample that is known to contain or suspected to contain the target polynucleotide. Alternatively, the invention may be carried out on a sample to confirm the identity of one or more target polynucleotides whose presence in the sample is known or expected. The sample may be a biological sample. The invention may be carried out in vitro on a sample obtained from or extracted from any organism or microorganism. The organism or microorganism is typically archaean, prokaryotic or eukaryotic and typically belongs to one the five kingdoms: plantae, animalia, fungi, monera and protista. The invention may be carried out in vitro on a sample obtained from or extracted from any virus. The sample is preferably a fluid sample. The sample typically comprises a body fluid of the patient. The sample may be urine, lymph, saliva, mucus or amniotic fluid but is preferably blood, plasma or serum. Typically, the sample is human in origin, but alternatively it may be from another mammal animal such as from commercially farmed animals such as horses, cattle, sheep or pigs or may alternatively be pets such as cats or dogs. Alternatively a sample of plant origin is typically obtained from a commercial crop, such as a cereal, legume, fruit or vegetable, for example wheat, barley, oats, canola, maize, soya, rice, bananas, apples, tomatoes, potatoes, grapes, tobacco, beans, lentils, sugar cane, cocoa, cotton. The sample may be a non-biological sample. The non-biological sample is preferably a fluid sample. Examples of a non-biological sample include surgical fluids, water such as drinking water, sea water or river water, and reagents for laboratory tests. The sample is typically processed prior to being assayed, for example by centrifugation or by passage through a membrane that filters out unwanted molecules or cells, such as red blood cells. The sample may be measured immediately upon being taken. The sample may also be typically stored prior to assay, preferably below −70° C. A transmembrane pore is a structure that permits hydrated ions driven by an applied potential to flow from one side of the membrane to the other side of the membrane. Any membrane may be used in accordance with the invention. Suitable membranes are well-known in the art. The membrane is preferably an amphiphilic layer. An amphiphilic layer is a layer formed from amphiphilic molecules, such as phospholipids, which have both hydrophilic and lipophilic properties. The amphiphilic layer may be a monolayer or a bilayer. The membrane is preferably a lipid bilayer. Lipid bilayers are models of cell membranes and serve as excellent platforms for a range of experimental studies. For example, lipid bilayers can be used for in vitro investigation of membrane proteins by single-channel recording. Alternatively, lipid bilayers can be used as biosensors to detect the presence of a range of substances. The lipid bilayer may be any lipid bilayer. Suitable lipid bilayers include, but are not limited to, a planar lipid bilayer, a supported bilayer or a liposome. The lipid bilayer is preferably a planar lipid bilayer. Suitable lipid bilayers are disclosed in International Application No. PCT/GB08/000563 (published as WO 2008/102121), International Application No. PCT/GB08/004127 (published as WO 2009/077734) and International Application No. PCT/GB2006/001057 (published as WO 2006/100484). Methods for forming lipid bilayers are known in the art. Suitable methods are disclosed in the Example. Lipid bilayers are commonly formed by the method of Montal and Mueller (Proc. Natl. Acad. Sci. USA., 1972; 69: 3561-3566), in which a lipid monolayer is carried on aqueous solution/air interface past either side of an aperture which is perpendicular to that interface. The method of Montal & Mueller is popular because it is a cost-effective and relatively straightforward method of forming good quality lipid bilayers that are suitable for protein pore insertion. Other common methods of bilayer formation include tip-dipping, painting bilayers and patch-clamping of liposome bilayers. In a preferred embodiment, the lipid bilayer is formed as described in International Application No. PCT/GB08/004127 (published as WO 2009/077734). In another preferred embodiment, the membrane is a solid state layer. A solid-state layer is not of biological origin. In other words, a solid state layer is not derived from or isolated from a biological environment such as an organism or cell, or a synthetically manufactured version of a biologically available structure. Solid state layers can be formed from both organic and inorganic materials including, but not limited to, microelectronic materials, insulating materials such as Si3N4, A1203, and SiO, organic and inorganic polymers such as polyamide, plastics such as Teflon® or elastomers such as two-component addition-cure silicone rubber, and glasses. The solid state layer may be formed from monatomic layers, such as graphene, or layers that are only a few atoms thick. Suitable graphene layers are disclosed in International Application No. PCT/US2008/010637 (published as WO 2009/035647). The method is typically carried out using (i) an artificial bilayer comprising a pore, (ii) an isolated, naturally-occurring lipid bilayer comprising a pore, or (iii) a cell having a pore inserted therein. The method is preferably carried out using an artificial lipid bilayer. The bilayer may comprise other transmembrane and/or intramembrane proteins as well as other molecules in addition to the pore. Suitable apparatus and conditions are discussed below. The method of the invention is typically carried out in vitro. The polynucleotide may be coupled to the membrane. This may be done using any known method. If the membrane is an amphiphilic layer, such as a lipid bilayer (as discussed in detail above), the polynucleotide is preferably coupled to the membrane via a polypeptide present in the membrane or a hydrophobic anchor present in the membrane. The hydrophobic anchor is preferably a lipid, fatty acid, sterol, carbon nanotube or amino acid. The polynucleotide may be coupled directly to the membrane. The polynucleotide is preferably coupled to the membrane via a linker. Preferred linkers include, but are not limited to, polymers, such as polynucleotides, polyethylene glycols (PEGs) and polypeptides. If a polynucleotide is coupled directly to the membrane, then some data will be lost as the characterising run cannot continue to the end of the polynucleotide due to the distance between the membrane and the helicase. If a linker is used, then the polynucleotide can be processed to completion. If a linker is used, the linker may be attached to the polynucleotide at any position. The linker is preferably attached to the polynucleotide at the tail polymer. The coupling may be stable or transient. For certain applications, the transient nature of the coupling is preferred. If a stable coupling molecule were attached directly to either the 5′ or 3′ end of a polynucleotide, then some data will be lost as the characterising run cannot continue to the end of the polynucleotide due to the distance between the bilayer and the helicase's active site. If the coupling is transient, then when the coupled end randomly becomes free of the bilayer, then the polynucleotide can be processed to completion. Chemical groups that form stable or transient links with the membrane are discussed in more detail below. The polynucleotide may be transiently coupled to an amphiphilic layer or lipid bilayer using cholesterol or a fatty acyl chain. Any fatty acyl chain having a length of from 6 to 30 carbon atoms, such as hexadecanoic acid, may be used. In preferred embodiments, polynucleotide is coupled to a lipid bilayer. Coupling of polynucleotides to synthetic lipid bilayers has been carried out previously with various different tethering strategies. These are summarised in Table 1 below. Polynucleotides may be functionalized using a modified phosphoramidite in the synthesis reaction, which is easily compatible for the addition of reactive groups, such as thiol, cholesterol, lipid and biotin groups. These different attachment chemistries give a suite of attachment options for polynucleotides. Each different modification group tethers the polynucleotide in a slightly different way and coupling is not always permanent so giving different dwell times for the polynucleotide to the bilayer. The advantages of transient coupling are discussed above. Coupling of polynucleotides can also be achieved by a number of other means provided that a reactive group can be added to the polynucleotide. The addition of reactive groups to either end of DNA has been reported previously. A thiol group can be added to the 5′ of ssDNA using polynucleotide kinase and ATPγS (Grant, G. P. and P. Z. Qin (2007). “A facile method for attaching nitroxide spin labels at the 5′ terminus of nucleic acids.” Alternatively, the reactive group could be considered to be the addition of a short piece of DNA complementary to one already coupled to the bilayer, so that attachment can be achieved via hybridisation. Ligation of short pieces of ssDNA have been reported using T4 RNA ligase I (Troutt, A. B., M. G. McHeyzer-Williams, et al. (1992). “Ligation-anchored PCR: a simple amplification technique with single-sided specificity.” A common technique for the amplification of sections of genomic DNA is using polymerase chain reaction (PCR). Here, using two synthetic oligonucleotide primers, a number of copies of the same section of DNA can be generated, where for each copy the 5′ of each strand in the duplex will be a synthetic polynucleotide. By using an antisense primer that has a reactive group, such as a cholesterol, thiol, biotin or lipid, each copy of the target DNA amplified will contain a reactive group for coupling. The transmembrane pore is preferably a transmembrane protein pore. A transmembrane protein pore is a polypeptide or a collection of polypeptides that permits hydrated ions, such as analyte, to flow from one side of a membrane to the other side of the membrane. In the present invention, the transmembrane protein pore is capable of forming a pore that permits hydrated ions driven by an applied potential to flow from one side of the membrane to the other. The transmembrane protein pore preferably permits analyte such as nucleotides to flow from one side of the membrane, such as a lipid bilayer, to the other. The transmembrane protein pore allows a polynucleotide, such as DNA or RNA, to be moved through the pore. The transmembrane protein pore may be a monomer or an oligomer. The pore is preferably made up of several repeating subunits, such as 6, 7 or 8 subunits. The pore is more preferably a heptameric or octameric pore. The transmembrane protein pore typically comprises a barrel or channel through which the ions may flow. The subunits of the pore typically surround a central axis and contribute strands to a transmembrane β barrel or channel or a transmembrane α-helix bundle or channel. The barrel or channel of the transmembrane protein pore typically comprises amino acids that facilitate interaction with analyte, such as nucleotides, polynucleotides or nucleic acids. These amino acids are preferably located near a constriction of the barrel or channel. The transmembrane protein pore typically comprises one or more positively charged amino acids, such as arginine, lysine or histidine, or aromatic amino acids, such as tyrosine or tryptophan. These amino acids typically facilitate the interaction between the pore and nucleotides, polynucleotides or nucleic acids. Transmembrane protein pores for use in accordance with the invention can be derived from β-barrel pores or α-helix bundle pores. β-barrel pores comprise a barrel or channel that is formed from β-strands. Suitable β-barrel pores include, but are not limited to, β-toxins, such as α-hemolysin, anthrax toxin and leukocidins, and outer membrane proteins/porins of bacteria, such as The transmembrane protein pore is preferably derived from Msp, preferably from MspA. Such a pore will be oligomeric and typically comprises 7, 8, 9 or 10 monomers derived from Msp. The pore may be a homo-oligomeric pore derived from Msp comprising identical monomers. Alternatively, the pore may be a hetero-oligomeric pore derived from Msp comprising at least one monomer that differs from the others. Preferably the pore is derived from MspA or a homolog or paralog thereof. A monomer derived from Msp comprises the sequence shown in SEQ ID NO: 2 or a variant thereof. SEQ ID NO: 2 is the MS-(B1)8 mutant of the MspA monomer. It includes the following mutations: D90N, D91N, D93N, D118R, D134R and E139K. A variant of SEQ ID NO: 2 is a polypeptide that has an amino acid sequence which varies from that of SEQ ID NO: 2 and which retains its ability to form a pore. The ability of a variant to form a pore can be assayed using any method known in the art. For instance, the variant may be inserted into a lipid bilayer along with other appropriate subunits and its ability to oligomerise to form a pore may be determined. Methods are known in the art for inserting subunits into membranes, such as lipid bilayers. For example, subunits may be suspended in a purified form in a solution containing a lipid bilayer such that it diffuses to the lipid bilayer and is inserted by binding to the lipid bilayer and assembling into a functional state. Alternatively, subunits may be directly inserted into the membrane using the “pick and place” method described in M. A. Holden, H. Bayley. J. Am. Chem. Soc. 2005, 127, 6502-6503 and International Application No. PCT/GB2006/001057 (published as WO 2006/100484). Over the entire length of the amino acid sequence of SEQ ID NO: 2, a variant will preferably be at least 50% homologous to that sequence based on amino acid identity. More preferably, the variant may be at least 55%, at least 60%, at least 65%, at least 70%, at least 75%, at least 80%, at least 85%, at least 90% and more preferably at least 95%, 97% or 99% homologous based on amino acid identity to the amino acid sequence of SEQ ID NO: 2 over the entire sequence. There may be at least 80%, for example at least 85%, 90% or 95%, amino acid identity over a stretch of 100 or more, for example 125, 150, 175 or 200 or more, contiguous amino acids (“hard homology”). Standard methods in the art may be used to determine homology. For example the UWGCG Package provides the BESTFIT program which can be used to calculate homology, for example used on its default settings (Devereux et al (1984) SEQ ID NO: 2 is the MS-(B1)8 mutant of the MspA monomer. The variant may comprise any of the mutations in the MspB, C or D monomers compared with MspA. The mature forms of MspB, C and D are shown in SEQ ID NOs: 5 to 7. In particular, the variant may comprise the following substitution present in MspB: A138P. The variant may comprise one or more of the following substitutions present in MspC: A96G, N102E and A138P. The variant may comprise one or more of the following mutations present in MspD: Deletion of G1, L2V, E5Q, L8V, D13G, W21A, D22E, K47T, I49H, I68V, D91G, A96Q, N102D, S103T, V104I, S136K and G141A. The variant may comprise combinations of one or more of the mutations and substitutions from Msp B, C and D. The variant preferably comprises the mutation L88N. The variant of SEQ ID NO: 2 has the mutation L88N in addition to all the mutations of MS-B1 and is called MS-B2. The pore used in the invention is preferably MS-(B2)8. Amino acid substitutions may be made to the amino acid sequence of SEQ ID NO: 2 in addition to those discussed above, for example up to 1, 2, 3, 4, 5, 10, 20 or 30 substitutions. Conservative substitutions replace amino acids with other amino acids of similar chemical structure, similar chemical properties or similar side-chain volume. The amino acids introduced may have similar polarity, hydrophilicity, hydrophobicity, basicity, acidity, neutrality or charge to the amino acids they replace. Alternatively, the conservative substitution may introduce another amino acid that is aromatic or aliphatic in the place of a pre-existing aromatic or aliphatic amino acid. Conservative amino acid changes are well-known in the art and may be selected in accordance with the properties of the 20 main amino acids as defined in Table 2 below. Where amino acids have similar polarity, this can also be determined by reference to the hydropathy scale for amino acid side chains in Table 3. One or more amino acid residues of the amino acid sequence of SEQ ID NO: 2 may additionally be deleted from the polypeptides described above. Up to 1, 2, 3, 4, 5, 10, 20 or 30 residues may be deleted, or more. Variants may include fragments of SEQ ID NO: 2. Such fragments retain pore forming activity. Fragments may be at least 50, 100, 150 or 200 amino acids in length. Such fragments may be used to produce the pores. A fragment preferably comprises the pore forming domain of SEQ ID NO: 2. Fragments must include one of residues 88, 90, 91, 105, 118 and 134 of SEQ ID NO: 2. Typically, fragments include all of residues 88, 90, 91, 105, 118 and 134 of SEQ ID NO: 2. One or more amino acids may be alternatively or additionally added to the polypeptides described above. An extension may be provided at the amino terminal or carboxy terminal of the amino acid sequence of SEQ ID NO: 2 or polypeptide variant or fragment thereof. The extension may be quite short, for example from 1 to 10 amino acids in length. Alternatively, the extension may be longer, for example up to 50 or 100 amino acids. A carrier protein may be fused to an amino acid sequence according to the invention. Other fusion proteins are discussed in more detail below. As discussed above, a variant is a polypeptide that has an amino acid sequence which varies from that of SEQ ID NO: 2 and which retains its ability to form a pore. A variant typically contains the regions of SEQ ID NO: 2 that are responsible for pore formation. The pore forming ability of Msp, which contains a β-barrel, is provided by β-sheets in each subunit. A variant of SEQ ID NO: 2 typically comprises the regions in SEQ ID NO: 2 that form β-sheets. One or more modifications can be made to the regions of SEQ ID NO: 2 that form β-sheets as long as the resulting variant retains its ability to form a pore. A variant of SEQ ID NO: 2 preferably includes one or more modifications, such as substitutions, additions or deletions, within its α-helices and/or loop regions. The monomers derived from Msp may be modified to assist their identification or purification, for example by the addition of histidine residues (a hist tag), aspartic acid residues (an asp tag), a streptavidin tag or a flag tag, or by the addition of a signal sequence to promote their secretion from a cell where the polypeptide does not naturally contain such a sequence. An alternative to introducing a genetic tag is to chemically react a tag onto a native or engineered position on the pore. An example of this would be to react a gel-shift reagent to a cysteine engineered on the outside of the pore. This has been demonstrated as a method for separating hemolysin hetero-oligomers (Chem Biol. 1997 July; 4(7):497-505). The monomer derived from Msp may be labelled with a revealing label. The revealing label may be any suitable label which allows the pore to be detected. Suitable labels include, but are not limited to, fluorescent molecules, radioisotopes, e.g.125I,35S, enzymes, antibodies, antigens, polynucleotides and ligands such as biotin. The monomer derived from Msp may also be produced using D-amino acids. For instance, the monomer derived from Msp may comprise a mixture of L-amino acids and D-amino acids. This is conventional in the art for producing such proteins or peptides. The monomer derived from Msp contains one or more specific modifications to facilitate nucleotide discrimination. The monomer derived from Msp may also contain other non-specific modifications as long as they do not interfere with pore formation. A number of non-specific side chain modifications are known in the art and may be made to the side chains of the monomer derived from Msp. Such modifications include, for example, reductive alkylation of amino acids by reaction with an aldehyde followed by reduction with NaBH4, amidination with methylacetimidate or acylation with acetic anhydride. The monomer derived from Msp can be produced using standard methods known in the art. The monomer derived from Msp may be made synthetically or by recombinant means. For example, the pore may be synthesized by in vitro translation and transcription (IVTT). Suitable methods for producing pores are discussed in International Application Nos. PCT/GB09/001690 (published as WO 2010/004273), PCT/GB09/001679 (published as WO 2010/004265) or PCT/GB10/000133 (published as WO 2010/086603). Methods for inserting pores into membranes are discussed. The transmembrane protein pore is also preferably derived from α-hemolysin (α-HL). The wild type α-HL pore is formed of seven identical monomers or subunits (i.e. it is heptameric). The sequence of one monomer or subunit of α-hemolysin-NN is shown in SEQ ID NO: 4. The transmembrane protein pore preferably comprises seven monomers each comprising the sequence shown in SEQ ID NO: 4 or a variant thereof. Amino acids 1, 7 to 21, 31 to 34, 45 to 51, 63 to 66, 72, 92 to 97, 104 to 111, 124 to 136, 149 to 153, 160 to 164, 173 to 206, 210 to 213, 217, 218, 223 to 228, 236 to 242, 262 to 265, 272 to 274, 287 to 290 and 294 of SEQ ID NO: 4 form loop regions. Residues 113 and 147 of SEQ ID NO: 4 form part of a constriction of the barrel or channel of α-HL. In such embodiments, a pore comprising seven proteins or monomers each comprising the sequence shown in SEQ ID NO: 4 or a variant thereof are preferably used in the method of the invention. The seven proteins may be the same (homoheptamer) or different (heteroheptamer). A variant of SEQ ID NO: 4 is a protein that has an amino acid sequence which varies from that of SEQ ID NO: 4 and which retains its pore forming ability. The ability of a variant to form a pore can be assayed using any method known in the art. For instance, the variant may be inserted into a lipid bilayer along with other appropriate subunits and its ability to oligomerise to form a pore may be determined. Methods are known in the art for inserting subunits into membranes, such as lipid bilayers. Suitable methods are discussed above. The variant may include modifications that facilitate covalent attachment to or interaction with the helicase. The variant preferably comprises one or more reactive cysteine residues that facilitate attachment to the helicase. For instance, the variant may include a cysteine at one or more of positions 8, 9, 17, 18, 19, 44, 45, 50, 51, 237, 239 and 287 and/or on the amino or carboxy terminus of SEQ ID NO: 4. Preferred variants comprise a substitution of the residue at position 8, 9, 17, 237, 239 and 287 of SEQ ID NO: 4 with cysteine (ABC, T9C, N17C, K237C, S239C or E287C). The variant is preferably any one of the variants described in International Application No. PCT/GB09/001690 (published as WO 2010/004273), PCT/GB09/001679 (published as WO 2010/004265) or PCT/GB10/000133 (published as WO 2010/086603). The variant may also include modifications that facilitate any interaction with nucleotides. The variant may be a naturally occurring variant which is expressed naturally by an organism, for instance by a Amino acid substitutions may be made to the amino acid sequence of SEQ ID NO: 4 in addition to those discussed above, for example up to 1, 2, 3, 4, 5, 10, 20 or 30 substitutions. Conservative substitutions may be made as discussed above. One or more amino acid residues of the amino acid sequence of SEQ ID NO: 4 may additionally be deleted from the polypeptides described above. Up to 1, 2, 3, 4, 5, 10, 20 or 30 residues may be deleted, or more. Variants may be fragments of SEQ ID NO: 4. Such fragments retain pore-forming activity. Fragments may be at least 50, 100, 200 or 250 amino acids in length. A fragment preferably comprises the pore-forming domain of SEQ ID NO: 4. Fragments typically include residues 119, 121, 135. 113 and 139 of SEQ ID NO: 4. One or more amino acids may be alternatively or additionally added to the polypeptides described above. An extension may be provided at the amino terminus or carboxy terminus of the amino acid sequence of SEQ ID NO: 4 or a variant or fragment thereof. The extension may be quite short, for example from 1 to 10 amino acids in length. Alternatively, the extension may be longer, for example up to 50 or 100 amino acids. A carrier protein may be fused to a pore or variant. As discussed above, a variant of SEQ ID NO: 4 is a subunit that has an amino acid sequence which varies from that of SEQ ID NO: 4 and which retains its ability to form a pore. A variant typically contains the regions of SEQ ID NO: 4 that are responsible for pore formation. The pore forming ability of α-HL, which contains a β-barrel, is provided by β-strands in each subunit. A variant of SEQ ID NO: 4 typically comprises the regions in SEQ ID NO: 4 that form β-strands. The amino acids of SEQ ID NO: 4 that form β-strands are discussed above. One or more modifications can be made to the regions of SEQ ID NO: 4 that form β-strands as long as the resulting variant retains its ability to form a pore. Specific modifications that can be made to the β-strand regions of SEQ ID NO: 4 are discussed above. A variant of SEQ ID NO: 4 preferably includes one or more modifications, such as substitutions, additions or deletions, within its α-helices and/or loop regions. Amino acids that form α-helices and loops are discussed above. The variant may be modified to assist its identification or purification as discussed above. Pores derived from α-HL can be made as discussed above with reference to pores derived from Msp. In some embodiments, the transmembrane protein pore is chemically modified. The pore can be chemically modified in any way and at any site. The transmembrane protein pore is preferably chemically modified by attachment of a molecule to one or more cysteines (cysteine linkage), attachment of a molecule to one or more lysines, attachment of a molecule to one or more non-natural amino acids, enzyme modification of an epitope or modification of a terminus. Suitable methods for carrying out such modifications are well-known in the art. The transmembrane protein pore may be chemically modified by the attachment of any molecule. For instance, the pore may be chemically modified by attachment of a dye or a fluorophore. Any number of the monomers in the pore may be chemically modified. One or more, such as 2, 3, 4, 5, 6, 7, 8, 9 or 10, of the monomers is preferably chemically modified as discussed above. The reactivity of cysteine residues may be enhanced by modification of the adjacent residues. For instance, the basic groups of flanking arginine, histidine or lysine residues will change the pKa of the cysteines thiol group to that of the more reactive S− group. The reactivity of cysteine residues may be protected by thiol protective groups such as dTNB. These may be reacted with one or more cysteine residues of the pore before a linker is attached. The molecule (with which the pore is chemically modified) may be attached directly to the pore or attached via a linker as disclosed in International Application Nos. PCT/GB09/001690 (published as WO 2010/004273), PCT/GB09/001679 (published as WO 2010/004265) or PCT/GB10/000133 (published as WO 2010/086603). Any Hel308 helicase may be used in accordance with the invention. Hel308 helicases are also known as ski2-like helicases and the two terms can be used interchangeably. The Hel308 helicase typically comprises the amino acid motif Q-X1-X2-G-R-A-G-R (hereinafter called the Hel308 motif; SEQ ID NO: 8). The Hel308 motif is typically part of the helicase motif VI (Tuteja and Tuteja, Eur. J. Biochem. 271, 1849-1863 (2004)). X1 may be C, M or L. X1 is preferably C. X2 may be any amino acid residue. X2 is typically a hydrophobic or neutral residue. X2 may be A, F, M, C, V, L, I, S, T, P or R. X2 is preferably A, F, M, C, V, L, I, S, T or P. X2 is more preferably A, M or L. X2 is most preferably A or M. The Hel308 helicase preferably comprises the motif Q-X1-X2-G-R-A-G-R-P (hereinafter called the extended Hel308 motif; SEQ ID NO: 9) wherein X1 and X2 are as described above. The most preferred Hel308 motifs and extended Hel308 motifs are shown in Table 5 below. The Hel308 helicase may comprise any of these preferred motifs. The Hel308 helicase is preferably one of the helicases shown in Table 4 below or a variant thereof. The Hel308 helicase is more preferably one of the helicases shown in Table 5 below or a variant thereof. The Hel308 helicase more preferably comprises the sequence of one of the helicases shown in Table 5, i.e. one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58, or a variant thereof. The Hel308 helicase more preferably comprises (a) the sequence of Hel308 Mbu (i.e. SEQ ID NO: 10) or a variant thereof, (b) the sequence of Hel308 Pfu (i.e. SEQ ID NO: 13) or a variant thereof, (c) the sequence of Hel308 Mok (i.e. SEQ ID NO: 29) or a variant thereof, (d) the sequence of Hel308 Mma (i.e. SEQ ID NO: 45) or a variant thereof, (e) the sequence of Hel308 Fac (i.e. SEQ ID NO: 48) or a variant thereof or (f) the sequence of Hel308 Mhu (i.e. SEQ ID NO: 52) or a variant thereof. The Hel308 helicase more preferably comprises the sequence shown in SEQ ID NO: 10 or a variant thereof. The Hel308 helicase more preferably comprises (a) the sequence of Hel308 Tga (i.e. SEQ ID NO: 33) or a variant thereof, (b) the sequence of Hel308 Csy (i.e. SEQ ID NO: 22) or a variant thereof or (c) the sequence of Hel308 Mhu (i.e. SEQ ID NO: 52) or a variant thereof. The Hel308 helicase most preferably comprises the sequence shown in SEQ ID NO: 33 or a variant thereof. A variant of a Hel308 helicase is an enzyme that has an amino acid sequence which varies from that of the wild-type helicase and which retains polynucleotide binding activity. In particular, a variant of any one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 is an enzyme that has an amino acid sequence which varies from that of any one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 and which retains polynucleotide binding activity. A variant of SEQ ID NO: 10 or 33 is an enzyme that has an amino acid sequence which varies from that of SEQ ID NO: 10 or 33 and which retains polynucleotide binding activity. The variant retains helicase activity. The variant must work in at least one of the two modes discussed below. Preferably, the variant works in both modes. The variant may include modifications that facilitate handling of the polynucleotide encoding the helicase and/or facilitate its activity at high salt concentrations and/or room temperature. Variants typically differ from the wild-type helicase in regions outside of the Hel308 motif or extended Hel308 motif discussed above. However, variants may include modifications within these motif(s). Over the entire length of the amino acid sequence of any one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58, such as SEQ ID NO: 10 or 33, a variant will preferably be at least 30% homologous to that sequence based on amino acid identity. More preferably, the variant polypeptide may be at least 40%, at least 45%, at least 50%, at least 55%, at least 60%, at least 65%, at least 70%, at least 75%, at least 80%, at least 85%, at least 90% and more preferably at least 95%, 97% or 99% homologous based on amino acid identity to the amino acid sequence of any one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58, such as SEQ ID NO: 10 or 33, over the entire sequence. There may be at least 70%, for example at least 80%, at least 85%, at least 90% or at least 95%, amino acid identity over a stretch of 150 or more, for example 200, 300, 400, 500, 600, 700, 800, 900 or 1000 or more, contiguous amino acids (“hard homology”). Homology is determined as described above. The variant may differ from the wild-type sequence in any of the ways discussed above with reference to SEQ ID NOs: 2 and 4. A variant of any one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 preferably comprises the Hel308 motif or extended Hel308 motif of the relevant wild-type sequence. For instance, a variant of SEQ ID NO: 10 preferably comprises the Hel308 motif of SEQ ID NO: 10 (QMAGRAGR; SEQ ID NO: 11) or extended Hel308 motif of SEQ ID NO: 10 (QMAGRAGRP; SEQ ID NO: 12). The Hel308 motif and extended Hel308 motif of each of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 are shown in Table 5. However, a variant of any one SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 may comprise the Hel308 motif or extended Hel308 motif from a different wild-type sequence. For instance, a variant of SEQ ID NO: 10 may comprise the Hel308 motif of SEQ ID NO: 13 (QMLGRAGR; SEQ ID NO: 14) or extended Hel308 motif of SEQ ID NO: 13 (QMLGRAGRP; SEQ ID NO: 15). A variant of any one SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 may comprise any one of the preferred motifs shown in Table 5. Variants of any one of SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55 and 58 may also include modifications within the Hel308 motif or extended Hel308 motif of the relevant wild-type sequence. Suitable modifications at X1 and X2 are discussed above when defining the two motifs. A variant of SEQ ID NO: 10 may lack the first 19 amino acids of SEQ ID NO: 10 and/or lack the last 33 amino acids of SEQ ID NO: 10. A variant of SEQ ID NO: 10 preferably comprises a sequence which is at least 70%, at least 75%, at least 80%, at least 85%, at least 90% or more preferably at least 95%, at least 97% or at least 99% homologous based on amino acid identity with amino acids 20 to 211 or 20 to 727 of SEQ ID NO: 10. The helicase may be covalently attached to the pore. The helicase is preferably not covalently attached to the pore. The application of a voltage to the pore and helicase typically results in the formation of a sensor that is capable of sequencing target polynucleotides. This is discussed in more detail below. Any of the proteins described herein, i.e. the transmembrane protein pores or Hel308 helicases, may be modified to assist their identification or purification, for example by the addition of histidine residues (a his tag), aspartic acid residues (an asp tag), a streptavidin tag, a flag tag, a SUMO tag, a GST tag or a MBP tag, or by the addition of a signal sequence to promote their secretion from a cell where the polypeptide does not naturally contain such a sequence. An alternative to introducing a genetic tag is to chemically react a tag onto a native or engineered position on the pore or helicase. An example of this would be to react a gel-shift reagent to a cysteine engineered on the outside of the pore. This has been demonstrated as a method for separating hemolysin hetero-oligomers (Chem Biol. 1997 July; 4(7):497-505). The pore and/or helicase may be labelled with a revealing label. The revealing label may be any suitable label which allows the pore to be detected. Suitable labels include, but are not limited to, fluorescent molecules, radioisotopes, e.g.125I,35S, enzymes, antibodies, antigens, polynucleotides and ligands such as biotin. Proteins may be made synthetically or by recombinant means. For example, the pore and/or helicase may be synthesized by in vitro translation and transcription (IVTT). The amino acid sequence of the pore and/or helicase may be modified to include non-naturally occurring amino acids or to increase the stability of the protein. When a protein is produced by synthetic means, such amino acids may be introduced during production. The pore and/or helicase may also be altered following either synthetic or recombinant production. The pore and/or helicase may also be produced using D-amino acids. For instance, the pore or helicase may comprise a mixture of L-amino acids and D-amino acids. This is conventional in the art for producing such proteins or peptides. The pore and/or helicase may also contain other non-specific modifications as long as they do not interfere with pore formation or helicase function. A number of non-specific side chain modifications are known in the art and may be made to the side chains of the protein(s). Such modifications include, for example, reductive alkylation of amino acids by reaction with an aldehyde followed by reduction with NaBH4, amidination with methylacetimidate or acylation with acetic anhydride. The pore and helicase can be produced using standard methods known in the art. Polynucleotide sequences encoding a pore or helicase may be derived and replicated using standard methods in the art. Polynucleotide sequences encoding a pore or helicase may be expressed in a bacterial host cell using standard techniques in the art. The pore and/or helicase may be produced in a cell by in situ expression of the polypeptide from a recombinant expression vector. The expression vector optionally carries an inducible promoter to control the expression of the polypeptide. These methods are described in described in Sambrook, J. and Russell, D. (2001). Molecular Cloning: A Laboratory Manual, 3rd Edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. The pore and/or helicase may be produced in large scale following purification by any protein liquid chromatography system from protein producing organisms or after recombinant expression. Typical protein liquid chromatography systems include FPLC, AKTA systems, the Bio-Cad system, the Bio-Rad BioLogic system and the Gilson HPLC system. The method of the invention involves measuring one or more characteristics of the target polynucleotide. The method may involve measuring two, three, four or five or more characteristics of the target polynucleotide. The one or more characteristics are preferably selected from (i) the length of the target polynucleotide, (ii) the identity of the target polynucleotide, (iii) the sequence of the target polynucleotide, (iv) the secondary structure of the target polynucleotide and (v) whether or not the target polynucleotide is modified. Any combination of (i) to (v) may be measured in accordance with the invention. For (i), the length of the polynucleotide may be measured using the number of interactions between the target polynucleotide and the pore. For (ii), the identity of the polynucleotide may be measured in a number of ways. The identity of the polynucleotide may be measured in conjunction with measurement of the sequence of the target polynucleotide or without measurement of the sequence of the target polynucleotide. The former is straightforward; the polynucleotide is sequenced and thereby identified. The latter may be done in several ways. For instance, the presence of a particular motif in the polynucleotide may be measured (without measuring the remaining sequence of the polynucleotide). Alternatively, the measurement of a particular electrical and/or optical signal in the method may identify the target polynucleotide as coming from a particular source. For (iii), the sequence of the polynucleotide can be determined as described previously. Suitable sequencing methods, particularly those using electrical measurements, are described in Stoddart D et al., Proc Natl Acad Sci, 12; 106(19):7702-7, Lieberman K R et al, J Am Chem Soc. 2010; 132(50):17961-72, and International Application WO 2000/28312. For (iv), the secondary structure may be measured in a variety of ways. For instance, if the method involves an electrical measurement, the secondary structure may be measured using a change in dwell time or a change in current flowing through the pore. This allows regions of single-stranded and double-stranded polynucleotide to be distinguished. For (v), the presence or absence of any modification may be measured. The method preferably comprises determining whether or not the target polynucleotide is modified by methylation, by oxidation, by damage, with one or more proteins or with one or more labels, tags or spacers. Specific modifications will result in specific interactions with the pore which can be measured using the methods described below. For instance, methylcyotsine may be distinguished from cytosine on the basis of the current flowing through the pore during its interation with each nucleotide. A variety of different types of measurements may be made. This includes without limitation: electrical measurements and optical measurements. Possible electrical measurements include: current measurements, impedance measurements, tunnelling measurements (Ivanov A P et al., Nano Lett. 2011 Jan. 12; 11(1):279-85), and FET measurements (International Application WO 2005/124888). Optical measurements may be combined 10 with electrical measurements (Soni G V et al., Rev Sci Instrum. 2010 January; 81(1):014301). The measurement may be a transmembrane current measurement such as measurement of ionic current flowing through the pore. Electrical measurements may be made using standard single channel recording equipment as describe in Stoddart D et al., Proc Natl Acad Sci, 12; 106(19):7702-7, Lieberman K R et al, J Am Chem Soc. 2010; 132(50):17961-72, and International Application WO-2000/28312. Alternatively, electrical measurements may be made using a multi-channel system, for example as described in International Application WO-2009/077734 and International Application WO-2011/067559. In a preferred embodiment, the method comprises: (a) contacting the target polynucleotide with a transmembrane pore and a Hel308 helicase such that the helicase controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore; and (b) measuring the current passing through the pore during one or more interactions to measure one or more characteristics of the target polynucleotide and thereby characterising the target polynucleotide. The methods may be carried out using any apparatus that is suitable for investigating a membrane/pore system in which a pore is inserted into a membrane. The method may be carried out using any apparatus that is suitable for transmembrane pore sensing. For example, the apparatus comprises a chamber comprising an aqueous solution and a barrier that separates the chamber into two sections. The barrier has an aperture in which the membrane containing the pore is formed. The methods may be carried out using the apparatus described in International Application No. PCT/GB08/000562 (WO 2008/102120). The methods may involve measuring the current passing through the pore during one or more interactions with the nucleotide(s). Therefore the apparatus may also comprise an electrical circuit capable of applying a potential and measuring an electrical signal across the membrane and pore. The methods may be carried out using a patch clamp or a voltage clamp. The methods preferably involve the use of a voltage clamp. The methods of the invention may involve the measuring of a current passing through the pore during one or more interactions with the nucleotide. Suitable conditions for measuring ionic currents through transmembrane protein pores are known in the art and disclosed in the Example. The method is typically carried out with a voltage applied across the membrane and pore. The voltage used is typically from +2 V to −2 V, typically −400 mV to +400 mV. The voltage used is preferably in a range having a lower limit selected from −400 mV, −300 mV, −200 mV, −150 mV, −100 mV, −50 mV, −20 mV and 0 mV and an upper limit independently selected from +10 mV, +20 mV, +50 mV, +100 mV, +150 mV, +200 mV, +300 mV and +400 mV. The voltage used is more preferably in the range 100 mV to 240 mV and most preferably in the range of 120 mV to 220 mV. It is possible to increase discrimination between different nucleotides by a pore by using an increased applied potential. The methods are typically carried out in the presence of any charge carriers, such as metal salts, for example alkali metal salt, halide salts, for example chloride salts, such as alkali metal chloride salt. Charge carriers may include ionic liquids or organic salts, for example tetramethyl ammonium chloride, trimethylphenyl ammonium chloride, phenyltrimethyl ammonium chloride, or 1-ethyl-3-methyl imidazolium chloride. In the exemplary apparatus discussed above, the salt is present in the aqueous solution in the chamber. Potassium chloride (KCl), sodium chloride (NaCl) or caesium chloride (CsCl) is typically used. KCl is preferred. The salt concentration may be at saturation. The salt concentration may be 3M or lower and is typically from 0.1 to 2.5 M, from 0.3 to 1.9 M, from 0.5 to 1.8 M, from 0.7 to 1.7 M, from 0.9 to 1.6 M or from 1 M to 1.4 M. The salt concentration is preferably from 150 mM to 1 M. As discussed above, Hel308 helicases surprisingly work under high salt concentrations. The method is preferably carried out using a salt concentration of at least 0.3 M, such as at least 0.4 M, at least 0.5 M, at least 0.6 M, at least 0.8 M, at least 1.0 M, at least 1.5 M, at least 2.0 M, at least 2.5 M or at least 3.0 M. High salt concentrations provide a high signal to noise ratio and allow for currents indicative of the presence of a nucleotide to be identified against the background of normal current fluctuations. The methods are typically carried out in the presence of a buffer. In the exemplary apparatus discussed above, the buffer is present in the aqueous solution in the chamber. Any buffer may be used in the method of the invention. Typically, the buffer is HEPES. Another suitable buffer is Tris-HCl buffer. The methods are typically carried out at a pH of from 4.0 to 12.0, from 4.5 to 10.0, from 5.0 to 9.0, from 5.5 to 8.8, from 6.0 to 8.7 or from 7.0 to 8.8 or 7.5 to 8.5. The pH used is preferably about 7.5. The methods may be carried out at from 0° C. to 100° C., from 15° C. to 95° C., from 16° C. to 90° C., from 17° C. to 85° C., from 18° C. to 80° C., 19° C. to 70° C., or from 20° C. to 60° C. The methods are typically carried out at room temperature. The methods are optionally carried out at a temperature that supports enzyme function, such as about 37° C. The method is typically carried out in the presence of free nucleotides or free nucleotide analogues and an enzyme cofactor that facilitate the action of the helicase. The free nucleotides may be one or more of any of the individual nucleotides discussed above. The free nucleotides include, but are not limited to, adenosine monophosphate (AMP), adenosine diphosphate (ADP), adenosine triphosphate (ATP), guanosine monophosphate (GMP), guanosine diphosphate (GDP), guanosine triphosphate (GTP), thymidine monophosphate (TMP), thymidine diphosphate (TDP), thymidine triphosphate (TTP), uridine monophosphate (UMP), uridine diphosphate (UDP), uridine triphosphate (UTP), cytidine monophosphate (CMP), cytidine diphosphate (CDP), cytidine triphosphate (CTP), cyclic adenosine monophosphate (cAMP), cyclic guanosine monophosphate (cGMP), deoxyadenosine monophosphate (dAMP), deoxyadenosine diphosphate (dADP), deoxyadenosine triphosphate (dATP), deoxyguanosine monophosphate (dGMP), deoxyguanosine diphosphate (dGDP), deoxyguanosine triphosphate (dGTP), deoxythymidine monophosphate (dTMP), deoxythymidine diphosphate (dTDP), deoxythymidine triphosphate (dTTP), deoxyuridine monophosphate (dUMP), deoxyuridine diphosphate (dUDP), deoxyuridine triphosphate (dUTP), deoxycytidine monophosphate (dCMP), deoxycytidine diphosphate (dCDP) and deoxycytidine triphosphate (dCTP). The free nucleotides are preferably selected from AMP, TMP, GMP, CMP, UMP, dAMP, dTMP, dGMP or dCMP. The free nucleotides are preferably adenosine triphosphate (ATP). The enzyme cofactor is a factor that allows the helicase to function. The enzyme cofactor is preferably a divalent metal cation. The divalent metal cation is preferably Mg2+, Mn2+, Ca2+ or Co2+. The enzyme cofactor is most preferably Mg2+. The target polynucleotide may be contacted with the Hel308 helicase and the pore in any order. In is preferred that, when the target polynucleotide is contacted with the Hel308 helicase and the pore, the target polynucleotide firstly forms a complex with the helicase. When the voltage is applied across the pore, the target polynucleotide/helicase complex then forms a complex with the pore and controls the movement of the polynucleotide through the pore. As discussed above, Hel308 helicases may work in two modes with respect to the nanopore. First, the method is preferably carried out using the Hel308 helicase such that it moves the target sequence through the pore with the field resulting from the applied voltage. In this mode the 3′ end of the DNA is first captured in the nanopore, and the enzyme moves the DNA into the nanopore such that the target sequence is passed through the nanopore with the field until it finally translocates through to the trans side of the bilayer. Alternatively, the method is preferably carried out such that the enzyme moves the target sequence through the pore against the field resulting from the applied voltage. In this mode the 5′ end of the DNA is first captured in the nanopore, and the enzyme moves the DNA through the nanopore such that the target sequence is pulled out of the nanopore against the applied field until finally ejected back to the cis side of the bilayer. The method of the invention most preferably involves a pore derived from MspA and a helicase comprising the sequence shown in SEQ ID NO: 8 or 10 or a variant thereof. Any of the embodiments discussed above with reference to MspA and SEQ ID NO: 8 and 10 may be used in combination. The invention also provides a method of forming a sensor for characterising a target polynucleotide. The method comprises forming a complex between a pore and a Hel308 helicase. The complex may be formed by contacting the pore and the helicase in the presence of the target polynucleotide and then applying a potential across the pore. The applied potential may be a chemical potential or a voltage potential as described above. Alternatively, the complex may be formed by covalently attaching the pore to the helicase. Methods for covalent attachment are known in the art and disclosed, for example, in International Application Nos. PCT/GB09/001679 (published as WO 2010/004265) and PCT/GB10/000133 (published as WO 2010/086603). The complex is a sensor for characterising the target polynucleotide. The method preferably comprises forming a complex between a pore derived from Msp and a Hel308 helicase. Any of the embodiments discussed above with reference to the method of the invention equally apply to this method. The present invention also provides kits for characterising a target polynucleotide. The kits comprise (a) a pore and (b) a Hel308 helicase. Any of the embodiments discussed above with reference to the method of the invention equally apply to the kits. The kit may further comprise the components of a membrane, such as the phospholipids needed to form a lipid bilayer. The kits of the invention may additionally comprise one or more other reagents or instruments which enable any of the embodiments mentioned above to be carried out. Such reagents or instruments include one or more of the following: suitable buffer(s) (aqueous solutions), means to obtain a sample from a subject (such as a vessel or an instrument comprising a needle), means to amplify and/or express polynucleotides, a membrane as defined above or voltage or patch clamp apparatus. Reagents may be present in the kit in a dry state such that a fluid sample resuspends the reagents. The kit may also, optionally, comprise instructions to enable the kit to be used in the method of the invention or details regarding which patients the method may be used for. The kit may, optionally, comprise nucleotides. The invention also provides an apparatus for characterising a target polynucleotide. The apparatus comprises a plurality of pores and a plurality of a Hel308 helicase. The apparatus preferably further comprises instructions for carrying out the method of the invention. The apparatus may be any conventional apparatus for polynucleotide analysis, such as an array or a chip. Any of the embodiments discussed above with reference to the methods of the invention are equally applicable to the apparatus of the invention. The apparatus is preferably set up to carry out the method of the invention. The apparatus preferably comprises: a sensor device that is capable of supporting the membrane and plurality of pores and being operable to perform polynucleotide characterising using the pores and helicases; at least one reservoir for holding material for performing the characterising; a fluidics system configured to controllably supply material from the at least one reservoir to the sensor device; and a plurality of containers for receiving respective samples, the fluidics system being configured to supply the samples selectively from the containers to the sensor device. The apparatus may be any of those described in International Application No. PCT/GB08/004127 (published as WO 2009/077734), PCT/GB10/000789 (published as WO 2010/122293), International Application No. PCT/GB10/002206 (not yet published) or International Application No. PCT/US99/25679 (published as WO 00/28312). Molecular motors are commonly used as a means for controlling the translocation of a polymer, particularly a polynucleotide, through a nanopore. Surprisingly, the inventors have found that molecular motors which are capable of binding to a target polynucleotide at an internal nucleotide, i.e. a position other than a 5′ or 3′ terminal nucleotide, can provide increased read lengths of the polynucleotide as the molecular motor controls the translocation of the polynucleotide through a nanopore. The ability to translocate an entire polynucleotide through a nanopore under the control of a molecular motor allows characteristics of the polynucleotide, such as its sequence, to be estimated with improved accuracy and speed over known methods. This becomes more important as strand lengths increase and molecular motors are required with improved processivity. The molecular motor used in the invention is particularly effective in controlling the translocation of target polynucleotides of 500 nucleotides or more, for example 1000 nucleotides, 5000, 10000 or 20000 or more. The invention thus provides a method of characterising a target polynucleotide, comprising: (a) contacting the target polynucleotide with a transmembrane pore and a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide such that the molecular motor controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore; and (b) measuring one or more characteristics of the target polynucleotide during one or more interactions and thereby characterising the target polynucleotide. Any of the embodiments discussed above in relation to the Hel308 methods of the invention equally apply to this method of the invention. A problem which occurs in sequencing polynucleotides, particularly those of 500 nucleotides or more, is that the molecular motor which is controlling translocation of the polynucleotide may disengage from the polynucleotide. This allows the polynucleotide to be pulled through the pore rapidly and in an uncontrolled manner in the direction of the applied field. Multiple instances of the molecular motor used in the invention bind to the polynucleotide at relatively short distances apart and thus the length of polynucleotide which can be pulled through the pore before a further molecular motor engages with the pore is relatively short. An internal nucleotide is a nucleotide which is not a terminal nucleotide in the target polynucleotide. For example, it is not a 3′ terminal nucleotide or a 5′ terminal nucleotide. All nucleotides in a circular polynucleotide are internal nucleotides. Generally, a molecular motor which is capable of binding at an internal nucleotide is also capable of binding at a terminal nucleotide, but the tendency for some molecular motors to bind at an internal nucleotide will be greater than others. For a molecular motor suitable for use in the invention, typically at least 10% of its binding to a polynucleotide will be at an internal nucleotide. Typically, at least 20%, at least 30%, at least 40% or at least 50% of its binding will be at an internal nucleotide. Binding at a terminal nucleotide may involve binding to both a terminal nucleotide and adjacent internal nucleotides at the same time. For the purposes of the invention, this is not binding to the target polynucleotide at an internal nucleotide. In other words, the molecular motor used in the invention is not only capable of binding to a terminal nucleotide in combination with one or more adjacent internal nucleotides. The molecular motor must be capable of binding to an internal nucleotide without concurrent binding to a terminal nucleotide. A molecular motor which is capable of binding at an internal nucleotide may bind to more than one internal nucleotide. Typically, the molecular motor binds to at least 2 internal nucleotides, for example at least 3, at least 4, at least 5, at least 10 or at least 15 internal nucleotides. Typically the molecular motor binds to at least 2 adjacent internal nucleotides, for example at least 3, at least 4, at least 5, at least 10 or at least 15 adjacent internal nucleotides. The at least 2 internal nucleotides may be adjacent or non-adjacent. The ability of a molecular motor to bind to a polynucleotide at an internal nucleotide may be determined by carrying out a comparative assay. The ability of a motor to bind to a control polynucleotide A is compared to the ability to bind to the same polynucleotide but with a blocking group attached at the terminal nucleotide (polynucleotide B). The blocking group prevents any binding at the terminal nucleotide of strand B, and thus allows only internal binding of a molecular motor. An example of this type of assay is disclosed in Example 4. Suitable molecular motors are well known in the art and typically include, but are not limited to, single and double strand translocases, such as polymerases, helicases, topoisomerases, ligases and nucleases, such as exonucleases. Preferably the molecular motor is a helicase, for example a Hel308 helicase. Examples of Hel308 helicases which are capable of binding at an internal nucleotide include, but are not limited to, Hel308 Tga, Hel308 Mhu and Hel308 Csy. Hence, the molecular motor preferably comprises (a) the sequence of Hel308 Tga (i.e. SEQ ID NO: 33) or a variant thereof or (b) the sequence of Hel308 Csy (i.e. SEQ ID NO: 22) or a variant thereof or (c) the sequence of Hel308 Mhu (i.e. SEQ ID NO: 52) or a variant thereof. The variant typically has at least 40% homology to SEQ ID NO: 33, 22 or 52 based on amino acid identity over the entire sequence and retains helicase activity. Further possible variants are discussed above. The molecular motor used in the invention may be made by any of the methods discussed above and may be modified or labelled as discussed above. The molecular motor may be used in the methods described herein or as part of the apparatus described herein. The invention further provides a method of forming a sensor for characterising a target polynucleotide, comprising forming a complex between a pore and a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide and thereby forming a sensor for characterising the target polynucleotide. The invention also provides use of a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide to control the movement of a target polynucleotide through a pore. The invention also provides a kit for characterising a target polynucleotide comprising (a) a pore and (b) a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide. The invention also provides an analysis apparatus for characterising target polynucleotides in a sample, comprising a plurality of pores and a plurality of a molecular motor which is capable of binding to the target polynucleotide at an internal nucleotide. The following Examples illustrate the invention. This Example illustrates the use of a Hel308 helicase (Hel308 MBu) to control the movement of intact DNA strands through a nanopore. The general method and substrate employed throughout this example is shown in Primers were designed to amplify a ˜400 bp fragment of PhiX174. Each of the 5′-ends of these primers included a 50 nucleotide non-complimentary region, either a homopolymeric stretch or repeating units of 10 nucleotide homopolymeric sections. These serve as identifiers for controlled translocation of the strand through a nanopore, as well as determining the directionality of translocation. In addition, the 5′-end of the forward primer was “capped” to include four 2′-O-Methyl-Uracil (mU) nucleotides and the 5′-end of the reverse primer was chemically phosphorylated. These primer modifications then allow for the controlled digestion of predominantly only the antisense strand, using lambda exonuclease. The mU capping protects the sense strand from nuclease digestion whilst the PO4 at the 5′ of the antisense strand promotes it. Therefore after incubation with lambda exonuclease only the sense strand of the duplex remains intact, now as single stranded DNA (ssDNA). The generated ssDNA was then PAGE purified as previously described. The DNA substrate design used in all the experiments described here is shown in Buffered solution: 400 mM-2 M KCl, 10 mM Hepes pH 8.0, 1 mM ATP, 1 mM MgCl2, 1 mM DTT
Electrical measurements were acquired from single MspA nanopores inserted in 1,2-diphytanoyl-glycero-3-phosphocholine lipid (Avanti Polar Lipids) bilayers. Bilayers were formed across ˜100 μm diameter apertures in 20 μm thick PTFE films (in custom Delrin chambers) via the Montal-Mueller technique, separating two 1 mL buffered solutions. All experiments were carried out in the stated buffered solution. Single-channel currents were measured on Axopatch 200B amplifiers (Molecular Devices) equipped with 1440A digitizers. Ag/AgCl electrodes were connected to the buffered solutions so that the cis compartment (to which both nanopore and enzyme/DNA are added) is connected to the ground of the Axopatch headstage, and the trans compartment is connected to the active electrode of the headstage. After achieving a single pore in the bilayer, DNA polynucleotide and helicase were added to 100 μL of buffer and pre-incubated for 5 mins (DNA=1.5 nM, Enzyme=1 μM). This pre-incubation mix was added to 900 μL of buffer in the cis compartment of the electrophysiology chamber to initiate capture of the helicase-DNA complexes in the MspA nanopore (to give final concentrations of DNA=0.15 nM, Enzyme=0.1 μM). Helicase ATPase activity was initiated as required by the addition of divalent metal (1 mM MgCl2) and NTP (1 mM ATP) to the cis compartment. Experiments were carried out at a constant potential of +180 mV. The addition of Helicase-DNA substrate to MspA nanopores as shown in For a given substrate, we observe a characteristic pattern of current transitions that reflects the DNA sequence (examples in In the implementation shown in Nanopore strand sequencing experiments of this type require ionic salts. The ionic salts are necessary to create a conductive solution for applying a voltage offset to capture and translocate DNA, and to measure the resulting sequence dependent current changes as the DNA passes through the nanopore. Since the measurement signal is dependent in the concentration of the ions, it is advantageous to use high concentration ionic salts to increase the magnitude of the acquired signal. For nanopore sequencing salt concentrations in excess of 100 mM KCl are ideal, and salt concentrations of 1 M KCl and above are preferred. However, many enzymes (including some helicases and DNA motor proteins) do not tolerate high salt conditions. Under high salt conditions the enzymes either unfold or lose structural integrity, or fail to function properly. The current literature for known and studied helicases shows that almost all helicases fail to function above salt concentrations of approximately 100 mM KCl/NaCl, and there are no reported helicases that show correct activity in conditions of 400 mM KCl and above. While potentially halophilic variants of similar enzymes from halotolerant species exist, they are extremely difficult to express and purify in standard expression systems (e.g. We surprisingly show in this Example that Hel308 from Mbu displays salt tolerance up to very high levels of KCl. We find that the enzyme retains functionality in salt concentrations of 400 mM KCl through to 2 M KCl, either in fluorescence experiments or in nanopore experiments ( Most helicases move along single-stranded polynucleotide substrates in uni-directional manner, moving a specific number of bases for each NTPase turned over. Although This Example illustrates the salt tolerance of a Hel308 helicase (Hel308 MBu) using a fluorescence assay for testing enzyme activity. A custom fluorescent substrate was used to assay the ability of the helicase to displace hybridised dsDNA ( Substrate DNA: 5′FAM-SEQ ID NO: 61 and SEQ ID NO: 62-BHQ1-3′. FAM=carboxyfluorescein and BHQ1=Black Hole Quencher-1 Capture DNA: SEQ ID NO: 62. The graph in In this Example, three different Hel308 helicases were used, namely Hel308 Mhu (SEQ ID NO: 52), Hel308 Mok (SEQ ID NO: 29) and Hel308 Mma (SEQ ID NO: 45). All experiments were carried out as previously described in Example 1 under the same experimental conditions (pore=MspA B2, DNA=400mer SEQ ID NO: 59 and 60, buffer=400 mM KCl, 10 mM Hepes pH 8.0, 1 mM dtt, 1 mM ATP, 0.1 mM MgCl2). The results are shown in This Example measures the internal binding capabilities of a number of Hel308 helicases using a fluorescence assay. Custom fluorescent substrates were used to assay the ability of the helicases to initiate on DNA lacking native 3′ ends, allowing them to subsequently displace hybridised dsDNA ( Substrate DNA: SEQ ID NO: 63 with a 5′ FAM; SEQ ID NO: 63 with a 5′ FAM and 3′ spacer ((spacer 9)4); SEQ ID NOs: 64 (with a 5′ FAM) and 65 separated by a spacer ((spacer 9)4); and SEQ ID NO: 62 with a 3′ BHQ1. Capture DNA: SEQ ID NO: 66. A number of different Hel308 helicase homologues were investigated for their mid-binding abilities, these included Hel308 Mbu, Hel308 Csy, Hel308 Tga, Hel308 Mma, Hel308 Mhu, Hel308 Min, Hel308 Mig, Hel308 Mmaz, Hel308 Mac, Hel308 Mok, Hel308 Mth, Hel308 Mba and Hel308 Mzh. The graph in This Example compares the use of two Hel308 helicases, Hel308 MBu and Hel 308 Tga, and their ability to control the movement of intact long DNA strands (900 mer) through a nanopore. The general method and substrate employed throughout this Example are shown in The DNA was formed by ligating a 50-polyT 5′ leader to a ˜900 base fragment of PhiX dsDNA. The leader also contains a complementary section to which SEQ ID NO: 69 with a Chol-tag was hybridized to allow the DNA to be tethered to the bilayer. Finally the 3′ end of the PhiX dsDNA was digested with AatII digestion enzyme to yield a 4 nt 3′-overhang of ACGT. Sequences used: SEQ ID NO: 67-900mer sense strand including 5′ leader and tether; SEQ ID NO: 68—anti-sense minus 4 base-pair leader 5′; and SEQ ID NO: 69 with several spacers and a Chol-tag at the 3′ end. Buffered solution: 400 mM-2 NaCl, 10 mM potassium ferrocyanide, 10 mM potassium ferricyanide, 100 mM Hepes, pH 8.0, 1 mM ATP, 1 mM MgCl2, Enzyme: Hel308 Mbu 1000 nM or Hel308 Tga 400 nM final. Electrical experiments were set up as described in Example 1 in order to achieve a single pore inserted into a lipid bilayer. After achieving a single pore in the bilayer, ATP (1 mM) and MgCl2 (1 mM) were added to the chamber. A control recording at +140 mV was run for 2 minutes. DNA polynucleotide SEQ ID NOs: 67, 68 and 69 (DNA=0.15 nM) were then added and DNA events observed. Finally, Hel308 helicase (Mbu 1000 nM or Tga, 400 nM) was added to the cis compartment of the electrophysiology chamber to initiate capture of the helicase-DNA complexes in the MspA nanopore. Experiments were carried out at a constant potential of +140 mV. The addition of Helicase-DNA substrate to MspA nanopores as shown in This Example illustrates that by employing the Hel308 helicase homologue Tga it is possible to control the translocation of a 5 kb strand of DNA. A similar experimental procedure was followed to that described in Example 5. It was observed that by employing the Hel308 Tga it was possible to detect the controlled translocation of an entire 5 kb strand of DNA through MS-(B1-G75S-G77S-L88N-Q126R)8. In an identical experiment using Hel308 Mbu, it was not possible to detect translocation of an entire 5 kB strand. This example compares the enzyme processivity of Hel308 Mbu helicase (SEQ ID NO: 10) with Hel308 Mok (SEQ ID NO: 29) using a fluorescence based assay. A custom fluorescent substrate was used to assay the ability of the helicase to displace hybridised dsDNA ( Additional custom fluorescent substrates were also used for control purposes. The substrate used as a negative control was identical to that of the one described in The invention relates to a new method of characterizing a target polynucleotide. The method uses a pore and a Hel308 helicase or amolecular motor which is capable of binding to the target polynucleotide at an internal nucleotide. The helicase or molecular motor controls the movement of the target polynucleotide through the pore. 1-43. (canceled) 44. A method of characterising a target polynucleotide, comprising:
(a) contacting the target polynucleotide with a transmembrane pore and a helicase which is capable of binding to the target polynucleotide at an internal nucleotide such that the helicase controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore; and (b) measuring one or more characteristics of the target polynucleotide during one or more interactions and thereby characterising the target polynucleotide. 45. A method according to 46. A method according to 47. A method according to 48. A method according to 49. A method according to (a) contacting the target polynucleotide with a transmembrane pore and a Hel308 helicase which is capable of binding to the target polynucleotide at an internal nucleotide such that the helicase controls the movement of the target polynucleotide through the pore and nucleotides in the target polynucleotide interact with the pore; and (b) measuring the current passing through the pore during one or more interactions to measure one or more characteristics of the target polynucleotide and thereby characterising the target polynucleotide. 50. A method according to 51. A method according to 52. A method according to 53. A method according to 54. A method according to 55. A method according to 56. A method according to 57. A method of forming a sensor for characterising a target polynucleotide, comprising forming a complex between a transmembrane pore and a helicase which is capable of binding to the target polynucleotide at an internal nucleotide and thereby forming a sensor for characterising the target polynucleotide. 58. Use of a helicase which is capable of binding to the target polynucleotide at an internal nucleotide to control the movement of a target polynucleotide through a transmembrane pore. 59. A kit for characterising a target polynucleotide comprising (a) a transmembrane pore and (b) a helicase which is capable of binding to the target polynucleotide at an internal nucleotide. 60. An analysis apparatus for characterising target polynucleotides in a sample, comprising a plurality of transmembrane pores and a plurality of helicases which are capable of binding to the target polynucleotide at an internal nucleotide.FIELD OF THE INVENTION
BACKGROUND OF THE INVENTION
SUMMARY OF THE INVENTION
DESCRIPTION OF THE FIGURES
DESCRIPTION OF THE SEQUENCE LISTING
DETAILED DESCRIPTION OF THE INVENTION
Hel308 Methods of the Invention
Thiol Stable Yoshina-Ishii, C. and S. G. Boxer (2003). “Arrays of mobile tethered vesicles on supported lipid bilayers.” Biotin Stable Nikolov, V., R. Lipowsky, et al. (2007). “Behavior of giant vesicles with anchored DNA molecules.” Cholestrol Transient Pfeiffer, I. and F. Hook (2004). “Bivalent cholesterol- based coupling of oligonucletides to lipid membrane assemblies.” Lipid Stable van Lengerich, B., R. J. Rawle, et al. “Covalent attachment of lipid vesicles to a fluid-supported bilayer allows observation of DNA-mediated vesicle interactions.” Ala aliphatic, hydrophobic, Met hydrophobic, neutral neutral Cys polar, hydrophobic, neutral Asn polar, hydrophilic, neutral Asp polar, hydrophilic, Pro hydrophobic, neutral charged (−) Glu polar, hydrophilic, Gln polar, hydrophilic, neutral charged (−) Phe aromatic, hydrophobic, Arg polar, hydrophilic, neutral charged (+) Gly aliphatic, neutral Ser polar, hydrophilic, neutral His aromatic, polar, hydrophilic, Thr polar, hydrophilic, neutral charged (+) Ile aliphatic, hydrophobic, Val aliphatic, hydrophobic, neutral neutral Lys polar, hydrophilic, Trp aromatic, hydrophobic, charged(+) neutral Leu aliphatic, hydrophobic, Tyr aromatic, polar, neutral hydrophobic Hydropathy scale Side Chain Hydropathy Ile 4.5 Val 4.2 Leu 3.8 Phe 2.8 Cys 2.5 Met 1.9 Ala 1.8 Gly −0.4 Thr −0.7 Ser −0.8 Trp −0.9 Tyr −1.3 Pro −1.6 His −3.2 Glu −3.5 Gln −3.5 Asp −3.5 Asn −3.5 Lys −3.9 Arg −4.5 Preferred Hel308 helicases Accession Description NP_578406.1 ski2-like helicase [ RecName: Full = Putative ski2-type helicase >pdb|2ZJ2|A Chain A, Archaeal Dna Helicase Hjm Apo State In Form 1 >pdb|2ZJ5|A Chain A, Archaeal Dna Helicase Hjm Complexed With Adp In Form 1 >pdb|2ZJ8|A Chain A, Archaeal Dna Helicase Hjm Apo State In Form 2 >pdb|2ZJA|A Chain A, Archaeal Dna Helicase Hjm Complexed With Amppcp In Form 2 >dbj|BAA32016.1| helicase [ 3638] NP_126564.1 ski2-like helicase [ RecName: Full = Putative ski2-type helicase >emb|CAB49795.1| DNA helicase [ NP_143168.1 ski2-like helicase [ RecName: Full = Putative ski2-type helicase >dbj|BAA30383.1| 715aa long hypothetical protein [ YP_004424773.1 ski2-like helicase [ [ YP_004623750.1 ski2-like helicase [ helicase [ YP_002307730.1 ski2-like helicase [ helicase [ YP_004763427.1 ski2-like helicase [ [ YP_002959236.1 ski2-like helicase [ type helicase, putative [ YP_004071709.1 ski2-type helicase [ ski2-type helicase [ YP_002994328.1 Putative ski2-type helicase [ Putative ski2-type helicase [ ZP_04875329.1 Type III restriction enzyme, res subunit family [ T469] >gb|EDY35111.1| Type III restriction enzyme, res subunit family [ YP_003436565.1 DEAD/DEAH box helicase [ DEAD/DEAH box helicase domain protein [ YP_004485304.1 ski2-type helicase [ helicase [ YP_004616424.1 DEAD/DEAH box helicase domain-containing protein [ DSM 4017] >gb|AEH61205.1| DEAD/DEAH box helicase domain protein [ ZP_04873370.1 Type III restriction enzyme, res subunit family [ T469] >ref|YP_003482774.1| DEAD/DEAH box helicase domain protein [ res subunit family [ DEAD/DEAH box helicase domain protein [ YP_004342552.1 ski2-type helicase [ helicase [ NP_071282.1 SKI2-family helicase [ 2P6R_A Chain A, Crystal Structure Of Superfamily 2 Helicase Hel308 In Complex With Unwound Dna >pdb|2P6U|A Chain A, Apo Structure Of The Hel308 Superfamily 2 Helicase YP_685308.1 ski2-like helicase [uncultured methanogenic archaeon RC-I] >sp|Q0W6L1.1|HELS_UNCMA RecName: Full = Putative ski2-type helicase >emb|CAJ35982.1| putative ski2-type helicase [uncultured methanogenic archaeon RC-I] YP_001048404.1 ski2-like helicase [ DEAD/DEAH box helicase domain protein [ YP_919908.1 DEAD/DEAH box helicase domain-containing protein [ Hrk 5] >gb|ABL77905.1| DEAD/DEAH box helicase domain protein [ YP_843229.1 ski2-like helicase [ DEAD/DEAH box helicase domain protein [ ZP_08045937.1 ski2-like helicase [ like helicase [ NP_280985.1 ski2-like helicase [ helicase [ RecName: Full = Putative ski2-type helicase >sp|B0R7Q2.1|HELS_HALS3 RecName: Full = Putative ski2-type helicase >gb|AAG20465.1| DNA repair protein [ [ YP_003357840.1 Holliday junction migration helicase [ SANAE] >dbj|BAI62857.1| Holliday junction migration helicase [ YP_003457479.1 DEAD/DEAH box helicase domain protein [ FS406-22] >gb|ADC68743.1| DEAD/DEAH box helicase domain protein [ YP_003127632.1 DEAD/DEAH box helicase domain protein [ AG86] >gb|ACV24132.1| DEAD/DEAH box helicase domain protein [ YP_003735335.1 ski2-like helicase [ helicase [ YP_503885.1 ski2-1ike helicase [ DEAD/DEAH box helicase-like protein [ BAJ48115.1 helicase [ [ [ YP_001405615.1 ski2-like helicase [ 6A8] >sp|A7IB61.1|HELS_METB6 RecName: Full = Putative ski2-type helicase >gb|ABS56972.1| DEAD/DEAH box helicase domain protein [ YP_306959.1 ski2-like helicase [ Fusaro] >sp|Q465R3.1|HELS_METBF RecName: Full = Putative ski2-type helicase >gb|AAZ72379.1| helicase [ YP_001031179.1 ski2-like helicase [ DEAD/DEAH box helicase domain protein [ YP_003541733.1 DEAD/DEAH box helicase [ 5219] >gb|ADE36088.1| DEAD/DEAH box helicase domain protein [ YP_004384692.1 putative Ski2-type helicase [ putative Ski2-type helicase [ YP_003725904.1 DEAD/DEAH box helicase domain-containing protein [ protein [ YP_003405271.1 DEAD/DEAH box helicase [ 5511] >gb|ADB62598.1| DEAD/DEAH box helicase domain protein [ YP_004244914.1 DEAD/DEAH box helicase [ 768-28] >gb|ADY01412.1| DEAD/DEAH box helicase domain protein [ YP_001540156.1 DEAD/DEAH box helicase domain-containing protein [ ski2-type helicase >gb|ABW01166.1| DEAD/DEAH box helicase domain protein [ NP_618094.1 ski2-like helicase [ C2A] >sp|Q8TL39.1|HELS_METAC RecName: Full = Putative ski2-type helicase >gb|AAM06574.1| helicase [ YP_003900980.1 DEAD/DEAH box helicase domain-containing protein [ DSM 14429] >gb|ADN49929.1| DEAD/DEAH box helicase domain protein [ YP_003896003.1 DEAD/DEAH box helicase domain-containing protein [ protein [ YP_003615773.1 DEAD/DEAH box helicase domain protein [ ME] >gb|ADG12809.1| DEAD/DEAH box helicase domain protein [ YP_183745.1 RNA helicase Ski2-like protein [ KOD1] >sp|Q5JGV6.1|HELS_PYRKO RecName: Full = Putative ski2-type helicase; Contains: RecName: Full = Endonuclease PI-PkoHel; AltName: Full = Pko Hel intein >dbj|BAD85521.1| RNA helicase Ski2 homolog [ YP_001322557.1 DEAD/DEAH box helicase domain-containing protein [ SB] >sp|A6UN73.1|HELS_METVS RecName: Full = Putative ski2-type helicase >gb|ABR53945.1| DEAD/DEAH box helicase domain protein [ YP_002467772.1 ski2-like helicase [ DEAD/DEAH box helicase domain protein [ YP_003480097.1 DEAD/DEAH box helicase [ DEAD/DEAH box helicase domain protein [ YP_004577043.1 ski2-type helicase [ ski2-type helicase [ YP_004742641.1 superfamily II helicase [ superfamily II helicase [ NP_632449.1 ski2-like helicase [ RecName: Full = Putative ski2-type helicase >gb|AAM30121.1| helicase [ YP_001097223.1 DEAD/DEAH box helicase domain-containing protein [ [ YP_004742247.1 DEAD/DEAH box helicase domain-containing protein [ protein [ YP_004794766.1 ski2-like helicase [ like helicase [ NP_988010.1 superfamily II helicase [ superfamily II helicase [ YP_565780.1 ski2-like helicase [ 6242] >sp|Q12WZ6.1|HELS_METBU RecName: Full = Putative ski2-type helicase >gb|ABE52030.1| DEAD/DEAH box helicase-like protein [ YP_001549808.1 DEAD/DEAH box helicase domain-containing protein [ [ YP_001548609.1 DEAD/DEAH box helicase domain-containing protein [ [ YP_001329359.1 DEAD/DEAH box helicase domain-containing protein [ [ YP_004595982.1 ski2-type helicase [ ski2-type helicase [ YP_656795.1 ski2-like helicase [ ATP-dependent DNA helicase [ CCC38992.1 ATP-dependent DNA helicase Hel308 [ YP_004035272.1 superfamily ii helicase [ 11551] >gb|ADQ65833.1| superfamily II helicase [ YP_137330.1 ski2-like helicase [ 43049] >sp|Q5UYM9.1|HELS_HALMA RecName: Full = Putative ski2-type helicase >gb|AAV47624.1| putative ski2-type helicase [ YP_001581577.1 DEAD/DEAH box helicase domain-containing protein [ [ EET90255.1 DEAD/DEAH box helicase domain protein [ NP_376477.1 helicase [ Full = Putative ski2-type helicase >dbj|BAK54341.1| Holliday junction migration helicase [ YP_001097792.1 DEAD/DEAH box helicase domain-containing protein [ [ ZP_08667240.1 DEAD/DEAH box helicase domain protein [ MY1] >gb|EGP92972.1| DEAD/DEAH box helicase domain protein [ sp. MY1] YP_254972.1 DNA helicase [ 639] >sp|Q4JC00.1|HELS_SULAC RecName: Full = Putative ski2-type helicase >gb|AAY79679.1| DNA helicase [ DSM 639] EFD92533.1 DEAD/DEAH box helicase domain protein [ YP_003176527.1 ski2-like helicase [ DEAD/DEAH box helicase domain protein [ EGD71904.1 DEAD/DEAH box helicase domain protein [ YP_001040230.1 DEAD/DEAH box helicase domain-containing protein [ F1] >gb|ABN69322.1| DEAD/DEAH box helicase domain protein [ ABZ07376.1 putative DEAD/DEAH box helicase [uncultured marine crenarchaeote HF4000_ANIW133M9] YP_001097458.1 DEAD/DEAH box helicase domain-containing protein [ [ ABZ08606.1 putative DEAD/DEAH box helicase [uncultured marine crenarchaeote HF4000_APKG3H9] YP_325906.1 ski2-like helicase [ 2160] >sp|Q31U46.1|HELS_NATPD RecName: Full = Putative ski2-type helicase >emb|CAI48337.1| ATP-dependent DNA helicase 1 [ YP_930665.1 DEAD/DEAH box helicase domain-containing protein [ DSM 4184] >gb|ABL88322.1| DEAD/DEAH box helicase domain protein [ YP_001435870.1 DEAD/DEAH box helicase [ DEAD/DEAH box helicase domain protein [ YP_003668634.1 DEAD/DEAH box helicase domain-containing protein [ protein [ ZP_08558598.1 ski2-like helicase [ helicase [ YP_002428409.1 DEAD/DEAH box helicase domain-containing protein [ protein [ YP_004336918.1 ATP-dependent, DNA binding helicase [ 768-20] >gb|AEA11606.1| ATP-dependent, DNA binding helicase [ ZP_08257442.1 DEAD/DEAH box helicase domain-containing protein [ domain-containing protein [ YP_004459284.1 DEAD/DEAH box helicase domain-containing protein [ W1] >gb|AEE94986.1| DEAD/DEAH box helicase domain protein [ NP_558924.1 ATP-dependent, DNA binding helicase [ IM2] >gb|AAL63106.1| ATP-dependent, DNA binding helicase [ YP_004409449.1 DEAD/DEAH box helicase domain-containing protein [ Ar-4] >gb|AEB94965.1| DEAD/DEAH box helicase domain-containing protein [ YP_003649556.1 DEAD/DEAH box helicase domain-containing protein [ protein [ ZP_06387115.1 DEAD/DEAH box helicase domain protein [ 98/2] >gb|ACX90562.1| DEAD/DEAH box helicase domain protein [ 2VA8_A Chain A, Dna Repair Helicase Hel308 >pdb|2VA8|B Chain B, Dna Repair Helicase Hel308 >emb|CAO85626.1| DNA helicase [ YP_004809267.1 ski2-type helicase [halophilic archaeon DL31] >gb|AEN06894.1| ski2-type helicase [halophilic archaeon DL31] ADX84345.1 DEAD/DEAH box helicase domain protein [ REY15A] >gb|ADX81629.1| DEAD/DEAH box helicase domain protein [ YP_002828439.1 DEAD/DEAH box helicase [ M.14.25] >ref|YP_002842325.1| DEAD/DEAH box helicase domain protein [ [ domain protein [ YP_002913571.1 DEAD/DEAH box helicase domain protein [ M.16.4] >gb|ACR40903.1| DEAD/DEAH box helicase domain protein [ Q97VY9.1 RecName: Full = Putative ski2-type helicase YP_002841682.1 DEAD/DEAH box helicase domain protein [ Y.N.15.51] >gb|ACP49760.1| DEAD/DEAH box helicase domain protein [ YP_002831080.1 DEAD/DEAH box helicase domain protein [ L.S.2.15] >ref|YP_003418425.1| DEAD/DEAH box helicase domain protein [ domain protein [ DEAD/DEAH box helicase domain protein [ YP_001054984.1 DEAD/DEAH box helicase domain-containing protein [ JCM 11548] >sp|A3MSA1.1|HELS_PYRCJ RecName: Full = Putative ski2-type helicase >gb|ABO07518.1| DEAD/DEAH box helicase domain protein [ NP_343811.1 DNA helicase related protein [ DEAD/DEAH box helicase [ DNA helicase related protein [ DEAD/DEAH box helicase domain protein [ YP_001152379.1 DEAD/DEAH box helicase domain-containing protein [ DSM 13514] >gb|ABP49727.1| DEAD/DEAH box helicase domain protein [ YP_001191456.1 DEAD/DEAH box helicase domain-containing protein [ DSM 5348] >gb|ABP95532.1| DEAD/DEAH box helicase domain protein [ NP_147034.2 holliday junction migration helicase [ K1] >sp|Q9YFQ8.2|HELS_AERPE RecName: Full = Putative ski2-type helicase >dbj|BAA79103.2| holliday junction migration helicase [ YP_024158.1 ski2-like helicase [ involved in UV-protection [ YP_003816358.1 Putative ski2-type helicase [ 345-15] >gb|ADL19327.1| Putative ski2-type helicase [ YP_003860265.1 DEAD/DEAH box helicase domain protein [ 17230] >gb|ADM28385.1| DEAD/DEAH box helicase domain protein [ NP_394295.1 ski2-like helicase [ 1728] >sp|Q9HJX7.1|HELS_THEAC RecName: Full = Putative ski2-type helicase >emb|CAC11964.1| DNA helicase related protein [ YP_876638.1 superfamily II helicase [ superfamily II helicase [ ZP_05571398.1 ski2-like helicase [ YP_004176252.1 DEAD/DEAH box helicase domain-containing protein [ DSM 2162] >gb|ADV64770.1| DEAD/DEAH box helicase domain protein [ YP_001737782.1 DEAD/DEAH box helicase domain-containing protein [ [ EGQ40435.1 superfamily II helicase [ YP_002567343.1 ski2-like helicase [ DEAD/DEAH box helicase domain protein [ YP_001793507.1 DEAD/DEAH box helicase domain-containing protein [ protein [ YP_003534088.1 ATP-dependent DNA helicase Hel308a [ DS2] >gb|ADE04048.1| ATP-dependent DNA helicase Hel308a [ YP_004037165.1 superfamily ii helicase [ 11551] >gb|ADQ67720.1| superfamily II helicase [ DSM 11551] NP_111333.1 ski2-like helicase [ GSS1] >sp|Q97AI2.1|HELS_THEVO RecName: Full = Putative ski2-type helicase >dbj|BAB59970.1| DNA helicase [ YP_002565871.1 DEAD/DEAH box helicase [ 49239] >gb|ACM56801.1| DEAD/DEAH box helicase domain protein [ CCC39675.1 ATP-dependent DNA helicase Hel308 [ YP_657401.1 ATP-dependent DNA helicase [ 16790] >emb|CAJ51759.1| ATP-dependent DNA helicase [ DSM 16790] YP_003535028.1 ATP-dependent DNA helicase Hel308b [ DS2] >gb|ADE02398.1| ATP-dependent DNA helicase Hel308b [ YP_003706863.1 DEAD/DEAH box helicase domain-containing protein [ A3] >gb|ADI35890.1| DEAD/DEAH box helicase domain protein [ ABD17736.1 helicase [ NP_613398.1 superfamily II helicase [ Predicted Superfamily II helicase [ CBH38575.1 putative ski2-type helicase [uncultured archaeon] EEZ93258.1 DEAD/DEAH box helicase domain protein [ EGQ40350.1 superfamily II helicase [ YP_004004246.1 dead/deah box helicase domain protein [ 2088] >gb|ADP77484.1| DEAD/DEAH box helicase domain protein [ YP_003850109.1 helicase [ predicted helicase [ YP_003424423.1 DEAD/DEAH box helicase domain-containing protein [ containing protein [ YP_004291107.1 DEAD/DEAH box helicase domain-containing protein [ AL-21] >gb|ADZ10135.1| DEAD/DEAH box helicase domain protein [ YP_447162.1 helicase [ helicase [ YP_004519549.1 DEAD/DEAH box helicase domain-containing protein [ SWAN-1] >gb|AEG17748.1| DEAD/DEAH box helicase domain protein [ NP_275949.1 DNA helicase related protein [ Delta H] >sp|O26901.1|HELS_METTH RecName: Full = Putative ski2-type helicase >gb|AAB85310.1| DNA helicase related protein [ ZP_05975717.2 putative Ski2-type helicase [ 2374] >gb|EFC93382.1| putative Ski2-type helicase [ ZP_03607647.1 hypothetical protein METSMIALI_00751 [ 2375] >gb|EEE41862.1| hypothetical protein METSMIALI_00751 [ YP_001273412.1 ATP-dependent helicase [ 35061] >gb|ABQ87044.1| ATP-dependent helicase [ ATCC 35061] YP_003247505.1 DEAD/DEAH box helicase domain protein [ M7] >gb|ACX73023.1| DEAD/DEAH box helicase domain protein [ NP_248116.1 SKI2 family helicase [ 2661] >sp|Q58524.1|HELS_METJA RecName: Full = Putative ski2-type helicase; Contains: RecName: Full = Endonuclease PI-MjaHel; AltName: Full = Mja Hel intein; AltName: Full = Mja Pep3 intein >gb|AAB99126.1| putative SKI2-family helicase [ YP_001324295.1 DEAD/DEAH box helicase domain-containing protein [ Nankai-3] >gb|ABR55683.1| DEAD/DEAH box helicase domain protein [ YP_003536960.1 Pre-mRNA splicing helicase [ mRNA splicing helicase [ YP_003131029.1 DEAD/DEAH box helicase domain protein [ 12940] >gb|ACV12296.1| DEAD/DEAH box helicase domain protein [ YP_002567151.1 DEAD/DEAH box helicase [ 49239] >gb|ACM58081.1| DEAD/DEAH box helicase domain protein [ YP_004035351.1 superfamily ii helicase [ 11551] >gb|ADQ65912.1| superfamily II helicase [ DSM 11551] YP_004808851.1 DEAD/DEAH box helicase domain-containing protein [halophilic archaeon DL31] >gb|AEN06478.1| DEAD/DEAH box helicase domain protein [halophilic archaeon DL31] XP_002716686.1 PREDICTED: DNA polymerase theta isoform 1 [ YP_656834.1 ATP-dependent DNA helicase [ 16790] >emb|CAJ51176.1| ATP-dependent DNA helicase [ DSM 16790] XP_003248103.1 PREDICTED: DNA polymerase theta-like isoform 1 [ ABC72356.1 ATP-dependent DNA helicase [ CCC39031.1 DEAD/DEAH box helicase [ XP_001165150.2 PREDICTED: DNA polymerase theta isoform 1 [ XP_003225852.1 PREDICTED: DNA polymerase theta-like [ XP_615375.3 PREDICTED: DNA polymerase theta [ PREDICTED: polymerase (DNA directed), theta-like [ theta-like [ XP_002813286.1 PREDICTED: LOW QUALITY PROTEIN: DNA polymerase theta-like [ AAR08421.2 DNA polymerase theta [ EAW79510.1 polymerase (DNA directed), theta, isoform CRA_a [ NP_955452.3 DNA polymerase theta [ RecName: Full = DNA polymerase theta; AltName: Full = DNA polymerase eta >gb|AAI72289.1| Polymerase (DNA directed), theta [synthetic polynucleotide] NP_001099348.1 DNA polymerase theta [ directed), theta (predicted), isoform CRA_a [ XP_003341262.1 PREDICTED: LOW QUALITY PROTEIN: DNA polymerase theta-like [ XP_001502374.3 PREDICTED: DNA polymerase theta [ XP_545125.3 PREDICTED: LOW QUALITY PROTEIN: DNA polymerase theta [ XP_002928855.1 PREDICTED: LOW QUALITY PROTEIN: DNA polymerase theta-like [ NP_084253.1 DNA polymerase theta isoform 1 [ polymerase theta [ theta, isoform CRA_a [ directed), theta [ theta [ AAK39635.1 DNA polymerase theta [ AAN39838.1 DNA polymerase Q [ XP_003412882.1 PREDICTED: DNA polymerase theta [ YP_003735206.1 DEAD/DEAH box helicase domain-containing protein [ B3] >gb|ADJ13414.1| DEAD/DEAH box helicase domain protein [ YP_004794841.1 pre-mRNA splicing helicase [ 33960] >gb|AEM55853.1| pre-mRNA splicing helicase [ XP_416549.2 PREDICTED: similar to DNA polymerase theta [ XP_003427319.1 PREDICTED: helicase POLQ-like isoform 2 [ XP_003202748.1 PREDICTED: DNA polymerase theta-like [ XP_969311.1 PREDICTED: similar to DNA polymerase theta [ TcasGA2_TC011380 [ ZP_08046037.1 DEAD/DEAH box helicase domain protein [ DX253] >gb|EFW90685.1| DEAD/DEAH box helicase domain protein [ YP_461714.1 helicase [ YP_003176510.1 DEAD/DEAH box helicase [ 12286] >gb|ACV46803.1] DEAD/DEAH box helicase domain protein [ YP_137400.1 Pre-mRNA splicing helicase [ 43049] >gb|AAV47694.1| Pre-mRNA splicing helicase [ ATCC 43049] NP_001184156.1 polymerase (DNA directed), theta [ NP_280861.1 pre-mRNA splicing helicase [ ATP-dependent DNA helicase [ pre-mRNA splicing helicase [ ATP-dependent DNA helicase [ YP_004595640.1 DEAD/DEAH box helicase domain-containing protein [ SH-6] >gb|AEH35761.1| DEAD/DEAH box helicase domain protein [ XP_001521144.2 PREDICTED: DNA polymerase theta, partial [ XP_003261953.1 PREDICTED: DNA polymerase theta, partial [ XP_001358456.2 GA19301 [ GA19301 [ ZP_08560003.1 DEAD/DEAH box helicase domain protein [ SARL4B] >gb|EGM34502.1| DEAD/DEAH box helicase domain protein [ XP_002187783.1 PREDICTED: similar to polymerase (DNA directed), theta [ XP_002112587.1 hypothetical protein TRIADDRAFT_25163 [ hypothetical protein TRIADDRAFT_25163 [ YP_003405139.1 DEAD/DEAH box helicase [ 5511] >gb|ADB62466.1| DEAD/DEAH box helicase domain protein [ EGV92665.1 DNA polymerase theta [ CBY24305.1 unnamed protein product [ YP_003130565.1 DEAD/DEAH box helicase domain protein [ 12940] >gb|ACV11832.1| DEAD/DEAH box helicase domain protein [ YP_003479811.1 DEAD/DEAH box helicase [ DEAD/DEAH box helicase domain protein [ EFB22383.1 hypothetical protein PANDA_000253 [ YP_003357334.1 putative ATP-dependent helicase [ SANAE] >dbj|BAI62351.1| putative ATP-dependent helicase [ SANAE] YP_325942.1 ATP-dependent DNA helicase 2 [ 2160] >emb|CAI48373.2| ATP-dependent DNA helicase 2 [ XP_002912509.1 PREDICTED: LOW QUALITY PROTEIN: helicase POLQ-like [ XP_002704678.1 PREDICTED: helicase, POLQ-like [ CAE47762.2 novel protein similar to humna DNA-directed polymerase theta (POLQ) [ XP_003205636.1 PREDICTED: helicase POLQ-like [ XP_544959.2 PREDICTED: helicase, POLQ-like [ EFX86757.1 hypothetical protein DAPPUDRAFT_312857 [ YP_003389641.1 DEAD/DEAH box helicase [ DEAD/DEAH box helicase domain protein [ XP_002602932.1 hypothetical protein BRAFLDRAFT_251779 [ BRAFLDRAFT_251779 [ YP_004144962.1 peptidase C14 caspase catalytic subunit p20 [ biserrulae WSM1271] >rel|YP_004614892.1| DEAD/DEAH box helicase domain-containing protein [ WSM2075] >gb|ADV14912.1| peptidase C14 caspase catalytic subunit p20 [ DEAD/DEAH box helicase domain protein [ WSM2075] XP_002124758.1 PREDICTED: similar to DNA polymerase theta [ XP_694437.5 PREDICTED: DNA polymerase theta [ XP_420565.1 PREDICTED: similar to DNA helicase HEL308 [ XP_003129397.1 PREDICTED: helicase POLQ-like [ EDL20278.1 mCG128467, isoform CRA_b [ XP_001517710.2 PREDICTED: helicase POLQ, partial [ AAH82601.1 Helicase, mus308-like ( XP_003384429.1 PREDICTED: DNA polymerase theta-like [ XP_003221282.1 PREDICTED: helicase POLQ-like [ NP_524333.1 mutagen-sensitive 308 [ [ AAX33507.1 LP14642p [ NP_001074576.1 helicase POLQ-like [ Full = Helicase POLQ-like; AltName: Full = Mus308-like helicase: AltName: Full = POLQ-like helicase >gb|AAI09171.2| Helicase, mus308-like ( [ YP_003523727.1 DEAD/DEAH box helicase domain protein [ ES-1] >gb|ADE11340.1| DEAD/DEAH box helicase domain protein [ XP_002120889.1 PREDICTED: similar to DNA helicase HEL308 [ XP_001892566.1 Type III restriction enzyme, res subunit family protein [ family protein [ ABZ09232.1 putative helicase conserved C-terminal domain protein [uncultured marine crenarchaeote HF4000_APKG7F11] XP_002814981.1 PREDICTED: LOW QUALITY PROTEIN: helicase POLQ-like [ XP_002717082.1 PREDICTED: DNA helicase HEL308 [ XP_001104832.1 PREDICTED: helicase, POLQ-like [ AAL85274.1 DNA helicase HEL308 [ NP_598375.2 helicase POLQ-like [ isoform CRA_a [ [synthetic polynucleotide] Q8TDG4.2 RecName: Full = Helicase POLQ-like; AltName: Full = Mus308-like helicase: AltName: Full = POLQ-like helicase XP_003265889.1 PREDICTED: helicase POLQ [ XP_002745688.1 PREDICTED: helicase POLQ-like [ XP_003310356.1 PREDICTED: LOW QUALITY PROTEIN: helicase POLQ-like [ NP_001014156.2 helicase, POLQ-like [ helicase, POLQ-like [ CRA_c [ XP_001850567.1 ATP-dependent DNA helicase MER3 [ ATP-dependent DNA helicase MER3 [ XP_003427318.1 PREDICTED: helicase POLQ-like isoform 1 [ XP_003143912.1 hypothetical protein LOAG_08332 [ protein LOAG_08332 [ CAG11187.1 unnamed protein product [ XP_001111254.2 PREDICTED: DNA polymerase theta isoform 2 [ XP_003414242.1 PREDICTED: helicase POLQ [ XP_002681870.1 predicted protein [ [ EAX05935.1 DNA helicase HEL308, isoform CRA_b [ AAH59917.1 Ascc3 protein [ ZP_07082808.1 DEAD/DEAH box helicase domain protein [ ATCC 33861] >gb|EFK55937.1| DEAD/DEAH box helicase domain protein [ XP_001494572.3 PREDICTED: LOW QUALITY PROTEIN: helicase POLQ-like [ XP_002714920.1 PREDICTED: activating signal cointegrator 1 complex subunit 3 [ XP_002598278.1 hypothetical protein BRAFLDRAFT_204526 [ BRAFLDRAFT_204526 [ XP_001943294.1 PREDICTED: helicase POLQ-like isoform 1 [ POLQ-like isoform 2 [ XP_002803889.1 PREDICTED: activating signal cointegrator 1 complex subunit 3-like [ XP_001651546.1 DNA polymerase theta [ [ CAA11679.1 RNA helicase [ XP_002837795.1 hypothetical protein [ protein product [ EGT47882.1 hypothetical protein CAEBREN_02542 [ EDL99655.1 activating signal cointegrator 1 complex subunit 3 (predicted), isoform CRA_b [ NP_932124.2 activating signal cointegrator 1 complex subunit 3 [ EDL05054.1 mCG119534 [ gi|352115865 DEAD/DEAH box helicase domain protein ZP_08963952.1 [ More preferred Hel308 helicases and most preferred Hel308 motifs and extended Hel308 motifs % Identity % Identity SEQ Hel308 Hel308 Extended ID NO: Helicase Names Pfu Mbu Hel308 motif Hel308 motif 10 Hel308 37% — QMAGRAGR QMAGRAGRP Mbu (SEQ ID NO: 11) (SEQ ID NO: 12) 13 Hel308 — 37% QMLGRAGR QMLGRAGRP Pfu (SEQ ID NO: 14) (SEQ ID NO: 15) DSM 3638 16 Hel308 34% 41% QMMGRAGR QMMGRAGRP Hvo (SEQ ID NO: 17) (SEQ ID NO: 18) 19 Hel308 35% 42% QMCGRAGR QMCGRAGRP Hla (SEQ ID NO: 20) (SEQ ID NO: 21) 22 Hel308 34% 34% QLCGRAGR QLCGRAGRP Csy (SEQ ID NO: 23) (SEQ ID NO: 24) 25 Hel308 35% 33% QMSGRAGR QMSGRAGRP Sso (SEQ ID NO: 26) (SEQ ID NO: 27) 28 Hel308 37% 44% QMAGRAGR QMAGRAGRP Mfr (SEQ ID NO: 11) (SEQ ID NO: 12) 29 Hel308 37% 34% QCIGRAGR QCIGRAGRP Mok (SEQ ID NO: 30) (SEQ ID NO: 31) 32 Hel308 40% 35% QCIGRAGR QCIGRAGRP Mig (SEQ ID NO: 30) (SEQ ID NO: 31) 33 Hel308 60% 38% QMMGRAGR QMMGRAGRP Tga (SEQ ID NO: 17) (SEQ ID NO: 18) EJ3 34 Hel308 57% 35% QMIGRAGR QMIGRAGRP Tba (SEQ ID NO: 35) (SEQ ID NO: 36) 37 Hel308 56% 35% QMMGRAGR QMMGRAGRP Tsi (SEQ ID NO: 17) (SEQ ID NO: 18) 38 Hel308 39% 60% QMAGRAGR QMAGRAGRP Mba (SEQ ID NO: 11) (SEQ ID NO: 12) 39 Hel308 38% 60% QMAGRAGR QMAGRAGRP Mac (SEQ ID NO: 11) (SEQ ID NO: 12) 40 Hel308 38% 60% QMAGRAGR QMAGRAGRP Mmah (SEQ ID NO: 11) (SEQ ID NO: 12) 41 Hel308 38% 60% QMAGRAGR QMAGRAGRP Mmaz (SEQ ID NO: 11) (SEQ ID NO: 12) 42 Hel308 39% 46% QMAGRAGR QMAGRAGRP Mth (SEQ ID NO: 11) (SEQ ID NO: 12) 43 Hel308 39% 57% QMAGRAGR QMAGRAGRP Mzh (SEQ ID NO: 11) (SEQ ID NO: 12) DSM 4017 44 Hel308 38% 61% QMAGRAGR QMAGRAGRP Mev (SEQ ID NO: 11) (SEQ ID NO: 12) Z-7303 45 Hel308 36% 32% QCIGRAGR QCIGRAGRP Mma (SEQ ID NO: 30) (SEQ ID NO: 31) 46 Hel308 37% 43% QMMGRAGR QMMGRAGRP Nma (SEQ ID NO: 17) (SEQ ID NO: 18) 47 Hel308 38% 45% QMAGRAGR QMAGRAGRP Mbo (SEQ ID NO: 11) (SEQ ID NO: 12) 48 Hel308 34% 32% QMIGRAGR QMIGRAGRP Fac (SEQ ID NO: 35) (SEQ ID NO: 36) 49 Hel308 40% 35% QCIGRAGR QCIGRAGRP Mfe (SEQ ID NO: 30) (SEQ ID NO: 31) 50 Hel308 24% 22% QCIGRAGR QCIGRAGRP Mja (SEQ ID NO: 30) (SEQ ID NO: 31) 51 Hel308 41% 33% QCIGRAGR QCIGRAGRP Min (SEQ ID NO: 30) (SEQ ID NO: 31) 52 Hel308 36% 40% QMAGRAGR QMAGRAGRP Mhu (SEQ ID NO: 11) (SEQ ID NO: 12) 53 Hel308 40% 40% QMAGRAGR QMAGRAGRP Afu (SEQ ID NO: 11) (SEQ ID NO: 12) DSM 4304 54 Hel308 35% 43% QMAGRAGR QMMGRAGRP Htu (SEQ ID NO: 11) (SEQ ID NO: 12) 55 Hel308 38% 45% QMFGRAGR QMFGRAGRP Hpa (SEQ ID NO: 56) (SEQ ID NO: 57) DX253 58 ski2-like 36.8% 42.0% QMFGRAGR QMFGRAGRP helicase NRC-1 (SEQ ID NO: 56) (SEQ ID NO: 57) Other Methods
Kits
Apparatus
Internally Binding Molecular Motors
Example 1
Materials and Methods
Nanopore:
Enzyme: Hel308 Mbu (ONLP3302, ˜7.7 μM) 12.5 μl->100 nM final.
Results and Discussion
Salt Tolerance
Effect of increasing salt concentration on pore current and DNA range Salt Open-pore current DNA range (KCl) (M) (pA) (pA) 0.4 180 25 1.0 440 55 2.0 840 75 Forward and Reverse Modes of Operation
Example 2
Example 3
Example 4
Example 5
Materials and Methods
Nanopore: MS-(B1-G75S-G77S-L88N-Q126R)8 (ONT Ref B2C)
Results and Discussion
Example 6
Example 7
SEQ ID NOs: 10, 13, 16, 19, 22, 25, 28, 29, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54 and 55 (Table 4) are aligned below The number below the * indicates the SEQ ID NO. The ″-″ are shown for alignment purposes only and do not form part of the sequences. * 1 95 10 Hel308 Mbu (1) --------------------------------MMIRELDIPRDIIGFYEDSGIKELYPPQAEAIEMGLLE-KKNLLAAIPTASGKTLLAELAMIK 53 Hel308 Afu (1) ------------------------------MKVEELAESISSYAVGILKEEGIEELFPPQAEAVEKVFS--GKNLLLAMPTAAGKTLLAEMAMVR 22 Hel308 Csy (1) --------------------------------MRISELDIPRPAIEFLEGEGYKKLYPPQAAAAKAGLTD-GKSVLVSAPTASGKTLIAAIAMIS 75 Hel308 Dth (1) MPGVDELLQQMGQGDLQGLSTVAVKEIPAREAEFSGIEGLPPPLKQALTESGIENFYTHQARAVNLVRK--GRSVVTATPTASGKSLIYNIPVLE 48 Hel308 Fac (1) ----------------------------------MKLSEITPSEFLKVTDNNDFTLYEHQEEAVAKLREN--KNVIVSVPTASGKTLIGYISIYD 19 Hel308 Hla (1) -------------------------------MQPSSLSGLPAGVGEALEAEGVAELYPPQEAAVEAGVAD-GESLVAAVPTASGKTLIAELAMLS 55 Hel308 Hpa (1) -------------------------------MNVADLTGLPDGVPEHFHAQGIEELYPPQAEAVEAGITE-GESVVASIPTASGKTFIAELAMLS 54 Hel308 Htu (1) -------------------------------MNLEELTGLPPGATDHFRGEGIEELYPPQADAVEAGATD-GENLVAAVPTASGKTMIAALSMLS 16 Hel308 Hvo (1) -------------------------------MRTADLTGLPTGIPEALRDEGIEELYPPQAEAVEAGLTD-GESLVAAVPTASGKTLIAELAMLS 39 Hel308 Mac (1) --------------------------------MKIESLDLPDEVKRFYENSGIPELYPPQAEAVEKGLLE-GKNLLAAIPTASGKTLLAELAMLK 38 Hel308 Mba (1) --------------------------------MKIESLDLPDEVKQFYLNSGIMELYPPQAEAVEKGLLE-GRNLLAAIPTASGKTLLAELAMLK 47 Hel308 Mbo (1) --------------------------------MQIQDLAIPEPLRQQYLGLGIRELYPPQAACVERGLLD-GKNLLVAIPTASGKTLIAEMAMHR 44 Hel308 Mev (1) --------------------------------METGKLELPEYVIQFYLDTGIEKLYPPQAEAVEKGLLD-NKNLLAAIPTASGKTLISELAMLK 49 Hel308 Mfe (1) --------------------------------------MPTNKILEILKDFGIEELRPPQKKALEKGLLDKNKNFLISIPTASGKTLIGEMALIN 28 Hel308 Mfr (1) ------------------------------------DLSLPKAFIQYYKDKGIESLYPPQSECIENGLLD-GADLLVAIPTASGKTLIAEMAMHA 52 Hel308 Mhu (1) --------------------------------MEIASLPLPDSFIRACHAKGIRSLYPPQAECIEKGLLE-GKNLLISIPTASGKTLLAEMAMWS 32 Hel308 Mig (1) -------------------------------------MQKYSHVFEVLKENGIKELRPPQKKVIEKGLLNKEKNFLICIPTASGKTLIGEMALIN 51 Hel308 Min (1) -------------------------------------------MDEILKFLGIKELRPPQKKALELGILDKKKNFLISIPTGAGKTVIAEMALIN 45 Hel308 Mma (1) -----------------------------------------MHVLDLLKENKITELRPPQKKVIDEGLFDKTKNFLICIPTASGKTLIGEMALLN 40 Hel308 Mmah (1) --------------------------------MKIEELDLPSEAIEVYLQAGIEELYPPQADAVEKGLLQ-GENLLAAIPTASGKTLLAEMAMLK 76 Hel308 Mmar (1) -------------------------------MDVADLPGVPEWLPDHLRDDGIEELYPPQAEAVEAGVTE-GENLVASIPTASGKTLIAELAMLS 41 Hel308 Mmaz (1) --------------------------------MKIESLDLPDEIKRFYENSGILELYPPQAEAVEKGLLE-GKNLLAAIPTASGKTLLAELAMLK 29 Hel308 Mok (1) ----------------------------------------MLMLMEVLKENGIAELRPPQKKVVEGGLLNKNKNFLICIPTASGKTLIGEMAFIN 42 Hel308 Mth (1) -----------------------------MLTIRDLIRWLPESVIELYEALGIDELYPPQAEAIERGLLD-GRNMIISVPTAAGKTLLAELAMLR 43 Hel308 Mzh (1) --------------------------------MNINNLNLPEKVKKYYTDTGIVDLYPPQREAVDKGLLD-GENIVAAIPTASGKTLLAELCMLK 46 Hel308 Nma (1) -------------------------------MNVEELSGLPPGARSHFQEQGIEELYPPQAEAVEAGATE-GENLVAAVPTASGKTMIAALSMLS 77 Hel308 Nth (1) --------------------------------MSETFYLLSERMQKKIWEMGWDEFTPVQDKTIPIVMNT-NKDVVVSSGTASGKTEAVFLPILS 13 Hel308 Pfu (1) -----------------------------MRVDELR---VDERIKSTLKERGIESFYPPQAEALKSGILE-GKNALISIPTASGKTLIAEIAMVH 25 Hel308 Sso (1) -------------------------MSLELEWMPIEDLKLPSNVIEIIKKRGIKKLNPPQTEAVKKGLLE-GNRLLLTSPTGSGKTLIAEMGIIS 34 Hel308 Tba (1) ---------MLSTKPKAYKRFSPIG--YAMQVDELSKFGVDERIIRKIKERGISEFYPPQAEALRSGVLN-GENLLLAIPTASGKTLVAEIVMLH 33 Hel308 Tga (1) -----------------------------MKVDELP---VDERLKAVLKERGIEELYPPQAEALKSGALE-GRNLVLAIPTASGKTLVSEIVMVN 37 Hel308 Tsi (1) ---------MKLNKLKSYINAFLLGMVMSMKVDELKSLGVDERILRLLRERGIEELYPPQADALKTEVLK-GKNLVLAIPTASGKTLVAEIVMIN 50 Hel308 Mja (1) ----------------------------------------MDKILEILKDFGIVELRPPQKKALERGLLDKNKNFLISIPTASGKTLIGEMALIN 78 Consensus (1) ---------------------------------------LP V L E GI ELYPPQAEAVE GLLD GKNLLIAIPTASGKTLIAELAML 96 190 Hel308 Mbu (63) AIREGG------KALYIVPLRALASEKFERFK-ELAP----FGIKVGISTGDLDSRADWLGVNDIIVATSEKTDSLLRNGTSWMD-------EIT Hel308 Afu (64) EAIKGG------KSLYVVPLRALAGEKYESFK-KWEK----IGLRIGISTGDYESRDEHLGDCDIIVTTSEKADSLIRNRASWIK-------AVS Hel308 Csy (63) HLSRNR-----GKAVYLSPLRALAAEKFAEFGKIGGIPL-GRPVRVGVSTGDFEKAGRSLGNNDILVLTNERMDSLIRRRPDWMD-------EVG Hel308 Dth (94) SIINDP----ASRALYLFPLKALTRDQLTSLEEFARLLAGKVHVDSAVYDGDTDPQARARIRSKPPNILLTNPDMLHRSFLPYHRSWQKFFSALK Hel308 Fac (60) TYLKGK------KSMYIVPLRSLAMEKFSELL-SLRN----LGVKVTMSIGDYDVPPSFVKNYDVIIATSERADSMLHRDPDILN-------YFG Hel308 Hla (64) SIERGG------KALYIVPLRALASEKKTEFE-RWEE----FGVTVGVSTGNYESDGEWLATRDIIVATSEKVDSLIRNGAPWID-------DLT Hel308 Hpa (64) SVARGG------KALYIVPLRALASEKKEEFE-EFEQ----YGVSIGVSTGNYESDGDWLASRDIIVATSEKVDSLVRNGAKWID-------DLS Hel308 Htu (64) AVQRGG------KALYIVPLRALASEKKEEFE-AYEE----FGVTTGVTTGNYESTDDWLATKDIIVATSEKVDSLVRNGADWLS-------ELT Hel308 Hvo (64) SVARGG------KALYIVPLRALASEKKAEFE-RWEE----YGIDVGVSTGNYESDGEWLSSRDIIVATSEKVDSLVRNNAAWMD-------QLT Hel308 Mac (63) SVLAGG------KALYIVPLRALASEKFRRFQ-DFSE----LGIRVGISTGDYDRRDEGLGINDIIVATSEKTDSLLRNETAWMQ-------EIS Hel308 Mba (63) SILAGG------KALYIVPLRALASEKFRRFR-EFSE----LGIRVGISTGDYDLRDEGLGVNDIIVATSEKTDSLLRNETVWMQ-------EIS Hel308 Mbo (63) HIANGG------KCLYIVPLKALASEKYEEFG-NK-------GVKVGLSTGDLDRRDDALGKNDIIVATSEKVDSLLRNGARWIP-------DIT Hel308 Mev (63) SISNGG------KCLYIVPLRALASEKFERFK-QFSS----IGVNIGISTGDFDSTDEWLGSNDIIVATSEKADSLLRNETSWMK-------DIT Hel308 Mfe (58) HLLDENKNPTNKKGIFIVPLKALASEKYEEFKNKYER----YGLRVALSIGDYD-EDEDLSRYHLIITTAEKLDSLWRHKIDWID-------DVS Hel308 Mfr (59) AIARGG------MCLYIVPLKALATEKAQEFK-GK-------GAEIGVATGDYDQKEKRLGSNDIVIATSEKVDSLLRNGVPWLS-------QVT Hel308 Mhu (63) RIAAGG------KCLYIVPLRALASEKYDEFS-KKG------VIRVGIATGDLDRTDAYLGENDIIVATSEKTDSLLRNRTPWLS-------QIT Hel308 Mig (59) HLLDENKTPTNKKGLFIVPLKALASEKYEEFKRKYEK----YGLKVALSIGDYD-EKEDLSSYNIIITTAEKLDSLMRHEIDWLN-------YVS Hel308 Min (53) HLLLDK----GKKGVYIVPLKALASEKYEEFKKKYEK----FGVRVALSIGDYD-EDEDLENYDLIITTAEKFDSLWRHGIKLS--------DIS Hel308 Mma (55) HILDENKNLTGKKGLFIVPLKALANEKFDEFREKYEK----YGIKVGLSIGDFD-TKENLSKFHIIITTSEKLDSLMRHNVEWIN-------DVS Hel308 Mmah (63) AIKKGG------KALYIVPLRALASEKFRDFK-RFES----LGIKTAISTGDFDSRDEWLGSNDIIVATSEKTDSLLRNSTPWMK-------DIT Hel308 Mmar (64) SVARGG------KALYIVPLRALASEKQADFE-EFEQ----YGLDIGVSTGNYESEGGWLADKDIVVATSEKVDSLVRNDAPWIE-------DLT Hel308 Mmaz (63) SVLNGG------KALYIVPLRALASEKFRRFQ-EFSV----LGMRVGISTGDYDRRDEGLGINDIIVATSEKTDSLLRNETAWMQ-------EIS Hel308 Mok (56) HLLDNNKTPTNKKGLFIVPLKALANEKYEEFKGKYEK----YGLKIALSIGDFD-EKEDLKGYDLIITTAEKLDSLIRHKVEWIK-------DIS Hel308 Mth (66) GALSGK------RSLYIVPLRALASEKFESFS-RFSK----LGLRVGISTGDFEKRDERLGRNDIIIATSEKADSLIRNGASWVR-------RIG Hel308 Mzh (63) SIGMGG------KCLYIVPLKALASEKYSRFR-EFES----LGIKVGIATGDLDSREEWLGKNDIIIATSEKVDSLLRNESSWMK-------EIN Hel308 Nma (64) AVQRGG------KALYIVPLRALASEKKAEFD-AYEE----FGVTTGVATGNYESTSEWLATKDIIVATSEKVDSLVRNGADWLS-------DLT Hel308 Nth (63) QIEKDAT--KDLKILYISPLKALINDQFERIIKLCEKSY-IPIHRWHGDVNQNKKKQLTKNPAGILQITPESIESLFINRTNELNYIL---SDIE Hel308 Pfu (63) RILTQG-----GKAVYIVPLKALAEEKFQEFQ-DWEK----IGLRVAMATGDYDSKDEWLGKYDIIIATAEKFDSLLRHGSSWIK-------DVK Hel308 Sso (70) FLLKNG-----GKAIYVTPLRALTNEKYLTFK-DWEL----IGFKVAMTSGDYDTDDAWLKNYDIIITTYEKLDSLWRHRPEWLN-------EVN Hel308 Tba (84) KLFTGG-----GKAVYLVPLKALAEEKYREFK-TWED----LGVRVAVTTGDYDSSEEWLGKYDIIIATSEKFDSLLRHKSRWIR-------DVT Hel308 Tga (63) KLIQEG-----GKAVYLVPLKALAEEKYREFK-EWEK----LGLKVAATTGDYDSTDDWLGRYDIIVATAEKFDSLLRHGARWIN-------DVK Hel308 Tsi (86) KILREG-----GKTVYLVPLKALAEEKYKEFK-FWEK----LGIRIAMTTGDYDSTEEWLGKYDIIIATSEKFDSLLRHKSPWIK-------DIN Hel308 Mja (56) HLLDGNKNPTNKKGIFIVPLKALASEKYEEFKSKYER----YGLRIALSIGDYD-EDEDLSKYHLIITTAEKLDSLWRHKIDWIN-------DVS Consensus (96) IL GG KALYIVPLRALASEKY EFK FE GVRVGISTGDYD DEWLG DIIVATSEKVDSLLRN WI DIT 191 285 Hel308 Mbu (140) TVVVDEIHLLDSKNRGPTLEVTITKLMRLNPD----VQVVALSATVGNAREMADWLG---AALVLSEWRPTDLHEGVLFGDAINFPG-SQKKIDR Hel308 Afu (141) CLVVDEIHLLDSEKRGATLEILVTKMRRMNKA----LRVIGLSATAPNVTEIAEWLD---ADYYVSDWRPVPLVEGVLCEGTLELFD----GAFS Hel308 Csy (145) LVIADEIHLIGDRSRGPTLEMVLTKLRGLRSS----PQVVALSATISNADEIAGWLD---CTLVHSTWRPVPLSEGVYQDGEVAMGDGSRHEVAA Hel308 Dth (185) YIVVDEVHTYRG-VMGSNMAWVFRRLRRICAQYGREPVFIFSSATIANPGQLCSALTGHEPEVIQKGGAPAGKKHFLLLDPEMQGAAQS------ Hel308 Fac (137) LVIIDEIHMISDPSRGPRLETVISSLLYLNPE----ILLLGLSATVSNIQEIAEWMN---AETVVSNFRAVPLETGIIFKGNLITDG-------- Hel308 Hla (141) CVVSDEVHLVDDPNRGPTLEVTLAKLRKVNPG----LQTVALSATVGNADVIAEWLD---AELVESDWRPIDLRMGVHFGNAIDFADGSKREVPV Hel308 Hpa (141) CVVADEVHLVNDAHRGPTLEVTLAKLRRVNPD----LQTVALSATVGNAGEMADWLD---ATLVDSTWRPIDLRKGVLYGQALHFDDGTQQELAR Hel308 Htu (141) CVVSDEVHLIDDRNRGPTLEVTLAKLRRLNPG----MQVVALSATVGNADEIADWLD---ASLVDTDWRPIDLQMGVHYGNALNFDDGSTREVPV Hel308 Hvo (141) CVVADEVHLVDDRHRGPTLEVTLAKLRRLNTN----LQVVALSATVGNAGVVSDWLD---AELVKSDWRPIDLKMGVHYGNAVSFADGSQREVPV Hel308 Mac (140) VVVVDEVHLIDSADRGPTLEVTLAKLRKMNPF----CQILALSATVGNADELAAWLD---AELVLSEWRPTDLMEGVFFDGTFFCKD-KEKLIEQ Hel308 Mba (140) VVVADEVHLIDSPDRGPTLEVTLAKLRKMNPS----CQILALSATVGNADELAVWLE---AELVVSEWRPTELLEGVFFNGTFYCKD-REKTVEQ Hel308 Mbo (137) LVVIDEIHLIDSPDRGPTLEMVIAKMRSKNPG----MQLIGLSATIGNPKVLAGWLD---AELVTSSWRPVDLRQGVFYDNRIQFAE-RMRPVKQ Hel308 Mev (140) TIVVDEIHLLDSADRGPTLEITIAKLLRLNPN----SQIIGLSATIGNAEEIAGWLD---AELVQSQWRPIELYEGVFLEDNINFKQ-SQKPIKN Hel308 Mfe (141) VVVVDEIHLINDESRGGTLEILLTKLKKFN------IQIIGLSATIGNPEELANWLN---AELIVDDWRPVELKKGIYKNGIIEFINGE-----N Hel308 Mfr (133) CLVVDEVHLIDDESRGPTLEMVITKLRHASPD----MQVIGLSATIGNPKELAGWLG---ADLITSDWRPVDLREGICYHNTIYFDN-EDKEIPA Hel308 Mhu (138) CIVLDEVHLIGSENRGATLEMVITKLRYTNPV----MQIIGLSATIGNPAQLAEWLD---ATLITSTWRPVDLRQGVYYNGKIRFSD-SERPIQG Hel308 Mig (142) VAIVDEIHMINDEKRGGTLEVLLTKLKNLD------VQIIGLSATIGNPEELAEWLN---AELIIDNWRPVKLRKGIFFQNKIMYLNGA-----C Hel308 Min (131) VVVVDEIHVIGDSERGGTLEVLLTKLKELD------VQIIGLSATIGNPEELSEWLN---AELLLDNWRPVELRKGIYREGVIEYLDGE------ Hel308 Mma (138) LAVIDEIHLIGDNERGGTLEVILTKLKNLN------AQIVGLSATIGNPEELSNWLN---AKLIVDGWRPVELKKGIYFENELEFLKNP-----A Hel308 Mmah (140) AVIVDEVHLLDSANRGPTLEVTLAKLKRLNPG----AQVVALSATVGNAMEIAQWLE---AKLVLSEWRPTYLHEGIFYGDAINFDE-DQTFIER Hel308 Mmar (141) CVVTDEVHLVDDGERGPTLEVTLAKLRRLNPD----LQTVALSATIGNAEALATWLD---AGLVDSDWRPIDLQKGVHYGQALHLEDGSQQRLSV Hel308 Mmaz (140) VVVADEVHLIDSPDRGPTLEITLSKLRRMNPS----CQVLALSATVGNADELAAWLD---AELVLSEWRPTDLMEGVFYNGIFYCKD-KEKPVGQ Hel308 Mok (139) VVVIDEIHLIGDESRGGTLEVLLTKLKTKKT-----IQIIGLSATIGNPEELAKWLN---AELIVDEWRPVKLKKGIGYGNKIMFIDDNGNTINE Hel308 Mth (143) VLVVDEIHLLDSANRGPTLEMTMTKLMHLNPE----MQVIGLSATIANGREIADWIK---GEIVSSDWRPVRLREGVLLEDRLVFPD-GEIQLEN Hel308 Mzh (140) TVVADEVHLLNSVNRGPTLEITLAKLIHLNPG----SQIIALSATIGNPEDIAGWLG---ARLVVSEWRPTDLYEGILLDGLLHIGN-IKKDIQD Hel308 Nma (141) CVVSDEVHLIDDRNRGPTLEVTLAKLRRLNPQ----LQVVALSATVGNADELADWLD---AELVDTDWRPIDLQMGVHYGNALNFDDGETREVPV Hel308 Nth (152) FIIIDELHAFLDNERGVHLRSLLSRLENYIKEK---PRYFALSATLNNFKLIKEWIN---YNDIKNVEIIDSNEDDKDLLLSLMHFDKGKDYKKP Hel308 Pfu (141) ILVADEIHLIGSRDRGATLEVILAHMLGKA-------QIIGLSATIGNPEELAEWLN---AELIVSDWRPVKLRRGVFYQGFVTWEDGSIDRFSS Hel308 Sso (148) YFVLDELHYLNDPERGPVVESVTIRAKRRN--------LLALSATISNYKQIAKWLG---AEPVATNWRPVPLIEGVIYPERKKKEYNVIFKDNT Hel308 Tba (162) LIVADEIHLLGSYDRGATLEMILSHMLGKA-------QILGLSATVGNAEELAEWLN---AKLVVSDWRPVKLRKGVFAHGQLIWEDGKVDKFPP Hel308 Tga (141) LVVADEVHLIGSYDRGATLEMILTHMLGRA-------QILALSATVGNAEELAEWLD---ASLVVSDWRPVQLRRGVFHLGTLIWEDGKVESYPE Hel308 Tsi (164) LVIADEIHLLGSYDRGATLEMILAHLDDKA-------QILGLSATVGNAEEVAEWLN---ADLVMSEWRPVALRKGVFYHGELFWEDGSIERFPT Hel308 Mja (139) VVVVDEIHLINDETRGGTLEILLTKLKEFN------VQIIGLSATIGNPDELAEWLN---AELIVDDWRPVELKKGIYKNEAIEFINGEIREIKA Consensus (191) VVVVDEIHLI D RGPTLEVLLAKLR LNP LQIIALSATIGNAEELAEWL AELVVSDWRPVDLR GVFY L F D I 286 380 Hel308 Mbu (227) LEK-----DDAVNLVLDTIKAEGQ-----CLVFESSRRNCAGFAKTASS---KVAKILDNDIMIKLAGIAEEVES--TGETDTAIVLANCIRKGV Hel308 Afu (225) TSRR----VKFEELVEECVAENGG-----VLVFESTRRGAEKTAVKLSA- -ITAKYVEN------EGLEKAILE--ENEGEMSRKLAECVRKGA Hel308 Csy (233) TGGG-----PAVDLAAESVAEGGQ-----SLIFADTRARSASLAAKASA---VIPEAKGADAAKLAAAAKKIISS--GGETKLAKTLAELVEKGA Hel308 Dth (273) ----------AIRVLQKALELGLR-----TIVYTQSRKMTELIAMWASQRAGRLKKYISAYRAGFLPEQRREIEQKLASGELLAVVSTSALELGI Hel308 Fac (217) -EKKHLGRDDEVSLIKESIESGGQ-----ALVFRNSRRNAEKYAQSMVN----------------FFDFQNDFEKLEIPPDLFNEAQANMVAHGV Hel308 Hla (229) ERGE----DQTARLVADALDTEEDGQGGSSLVFVNSRRNAESSARKLTD---VTGPRLTDDERDQLRELADEIRS--GSDTDTASDLADAVEQGS Hel308 Hpa (229) -GNE----KETAALVRDTLEDGGS-----SLVFVNSRRNAEAAAKRLAD---VTKTHLTDDERRDLLDIADQIRD--VSDTETSDDLATAIEKGA Hel308 Htu (229) EGSE----KQEAALVRDILREGGS-----SLVFVNSRRNAEGAAKRLGQ---VSSREITEDERAELAELADDIRD--DSDTETSADLADCVERGA Hel308 Hvo (229) GRGE----RQTPALVADALEGDGEGDQGSSLVFVNSRRNAESAARRMAD---VTERYVTGDERSDLAELAAEIRD--VSDTETSDDLANAVAKGA Hel308 Mac (227) PTK-----DEAINLVLDTLREGGQ-----CLVFESSRKNCMGFAKKATS---AVKKTLSAEDKEKLAGIADEILE--NSETDTASVLASCVRAGT Hel308 Mba (227) STK-----DEAVNLALDTLKKDGQ-----CLVFESSRKNCMAFAKKAAS---TVKKTLSAEDRNALAGIADEILE--NSETDTSTNLAVCIRSGT Hel308 Mbo (224) VSKN----YDDLNLCLDTIAEGGQ-----CLVFVSSRRNAEAFAKRAAG---AIKSEDA-----ALAACAERLLE--GTPTEMVKTLAACVAKGA Hel308 Mev (227) IVK-----DTAVNLVLDTIDENGQ-----CLVFESSRRNCAGFAKKAKS---KVGKSLDKGLLAELNNIAEEVLE--TSDTETTKELASCIKRGT Hel308 Mfe (222) REIKAINNNDIYNLVVDCVKDGGC-----CIVFCNTKRGAVNEAKKLN-----LKKFLTNEEKRKLKEVAEEILSILEPPTEMCKTLAECILNGS Hel308 Mfr (220) PAK-----TEDINLLLDCVADGGQ-----CLVFVSSRRNAEGYAKRAAT---ALKCSHA-----ALDSIAEKLEA--AAETDMGRVLATCVKKGA Hel308 Mhu (225) KTK-----HDDLNLCLDTIEEGGQ-----CLVFVSSRRNAEGFAKKAAG---ALKAGSP-----DSKALAQELRR--LRDRDEGNVLADCVERGA Hel308 Mig (223) KELPNFSNNPMLNLVLDCVKEGGC-----CLVFCNSKNGAVSEAKKLN-----LKKYLSNSEKYELQKLKEEILSILDPPTETCKTLAECLEKGV Hel308 Min (211) -------VKECQDIVKEVVKDNGS-----VIIFCPTKKKAENRALSLD-----LSDLLKKSEKRKLEEISEELLSLFDPPTELCKKLASCVRKGI Hel308 Mma (219) KKIKQVSRNNLTDLIVDSVEEKGS-----CLIFCNSKRNAVGEAKKHN-----LAKYLTRTEQHELNKLSEEILSILDRPVETCKALSKCIQNGV Hel308 Mmah (227) RHK-----EDSVNLVIDTVIQGGQ-----CLVFDSSRRNCVGFAKKCAP---AVGELLDRQNRNELEEVAKEVLE--NGETKLTETLAYCIKKGV Hel308 Mmar (229) QNNE----KQTAAIVRDTLEDDGS-----TLVFVNSRRNAEAAAGRLAN---TVRPHLSTEERDQLADIAEEIRD--VSDTETSDDLADAVADGA Hel308 Mmaz (227) PTK-----DEAVNLVLDTIKEGGQ-----CLVFESSRKNCMGFAKKAVS---AVKKTLSNEDRETLAGIADEIIE--NSETDVSSVLATCVRSGT Hel308 Mok (226) VIVDEISKNNMFNLVVDSILKDGS-----CIIFCNSKRGAVGEAKKLN-----LKKYLSPDEISELRHLKEEVLSVLDNPTKTCKDLAECIEKGV Hel308 Mth (230) RNR-----DPVLNLVLDTVDQGGQ-----MLIFESTRRNAESMAKKVSG---ALQESGE------TIELAERLS----GEGKTAKKLAMCLRHGA Hel308 Mzh (227) ESR-----DDAVNLVIDTVKDKGQ-----CLVFESSRRNCMGFAKKAGK---WVSKILDEHDTIQLKSLSQEIGE--AGETEIADVLSRCVRQGV Hel308 Nma (229) EAGE----KQEAALVRDILQEGGS-----SLVFVNSRRNAEAAARRLGQ---VSSRELTAGEQNDLAALATEIRE--DSDTETSQDLADCVERGA Hel308 Nth (241) ----------ID-LYQDLRELTKN---VHSLIFCNSRAEVEETTLYLNR---LANREVNTELYLAHHSSIDKKER-EYVEKTMANSKSPKSVVTT Hel308 Pfu (226) ----------WEELVYDAIRKKKG-----ALIFVNMRRKAERVALELSK---KVKSLLTKPEIRALNELADSLE-----ENPTNEKLAKAIRGGV Hel308 Sso (232) TKKVHG-DDAIIAYTLDSLSKNGQ-----VLVFRNSRKMAESTALKIAN---YMNFVSLDEN--ALSEILKQLDDIEEGGSDEKELLKSLISKGV Hel308 Tba (247) Q---------WDSLVIDAVKKGKQ-----ALVFVNTRRSAEKEAGMLGK---KVRRLLTKPEARRLKELAESLE-----SNPTNDKLKEVLVNGA Hel308 Tga (226) N---------WYSLVVDAVKRGKG-----ALVFVNTRRSAEKEALALSK---LVSSHLTKPEKRALESLASQLE-----DNPTSEKLKRALRGGV Hel308 Tsi (249) Q---------WDSLVIDALKKGKQ-----ALVFVNTRRSAEKEALLLAG---KIQRFLTKPEERKLKQLADGLD-----TTPTNQKLKEALTKGV Hel308 Mja (225) VDN-----NDIYNLVVDCVKEGGC-----CLVFCNTKRNAVNEAKKLN-----LKKFLTEEEKIRLKEIAEEILSILEPPTEMCKTLAECILNGS Consensus (286) LVLDTV EGGQ LVF NSRRNAE AKKLA V K LT E L LAEEI ETETS LA CV KG 381 475 Hel308 Mbu (307) AFHHAGLNSNH------------RKLVENGFRQNLIKVISSTPTLAA------------------------------------------------ Hel308 Afu (300) AFHHAGLLNGQ------------RRVVEDAFRRGNIKVVVATPTLAA------------------------------------------------ Hel308 Csy (313) AFHHAGLNQDC------------RSVVEEEFRSGRIRLLASTPTLAA------------------------------------------------ Hel308 Dth (353) DIGHLDLCLLVGYPGSVMATMQRGGRVGRSGRDSAIMLIGHEDALDQYLLRNPREFFSLEPESAVINPDNPSIMRRHLVCAAAEKPIALQEMMLD Hel308 Fac (290) MFHHAGLSNDQ------------RTMIEKLFKQGYIKILTATPTLAA------------------------------------------------ Hel308 Hla (315) AFHHAGLRSED------------RARVEDAFRDRLIKCISATPTLAA------------------------------------------------ Hel308 Hpa (309) AFHHAGLASDH------------RSLVEDAFRDKLIKVISATPTLAA------------------------------------------------ Hel308 Htu (310) AFHHAGLSSTQ------------RSLVEDAFRDRLLKVISATPTLAA------------------------------------------------ Hel308 Hvo (315) AFHHAGLAAEH------------RTLVEDAFRDRLIKCICATPTLAA------------------------------------------------ Hel308 Mac (307) AFHHAGLTSPL------------RELVETGFREGYVKLISSTPTLAA------------------------------------------------ Hel308 Mba (307) AFHHAGLTTPL------------RELVEDGFRAGRIKLISSTPTLAA------------------------------------------------ Hel308 Mbo (300) AFHHAGLSRKE------------RSIVEEAFRKNLLKCISSTPTLAA------------------------------------------------ Hel308 Mev (307) AFHHAGLNSAQ------------RKIVEDNFRNNKIKVISSTPTLAA------------------------------------------------ Hel308 Mfe (307) AFHHAGLTYQH------------RKIVEDAFRNKLIKVICCTPTLSV------------------------------------------------ Hel308 Mfr (295) AFHHAGMNRMQ------------RTLVEGGFRDGFIKSISSTPTLAA------------------------------------------------ Hel308 Mhu (300) AFHHAGLIRQE------------RTIIEEGFRNGYIEVIAATPTLAA------------------------------------------------ Hel308 Mig (308) AFHHAGLTYEH------------RKIVEEGFRNKLIKVICCTPTLSA------------------------------------------------ Hel308 Min (289) AFHHSGLTYEH------------RKIIEKAFRERILKVICSTTTLAF------------------------------------------------ Hel308 Mma (304) AFHHAGLTYKH------------RKIVEDGFRNRLIKVICCTPTLSA------------------------------------------------ Hel308 Mmah (307) AFHHAGLNSAH------------RRIVEDAFRNNLIKMICSTPTLAA------------------------------------------------ Hel308 Mmar (310) AFHHAGLSRGH------------RELVEDAFRDRLVKVVCATPTLAA------------------------------------------------ Hel308 Mmaz (307) AFHHAGLTTPL------------RELVENGFREGRIKIISSTPTLAA------------------------------------------------ Hel308 Mok (311) AFHHAGLTYEQ------------RKIVEEGFRKKLIKAICCTPTLSA------------------------------------------------ Hel308 Mth (302) AFHHAGLLPEQ------------RRLIELGFRQNVVKVIACTPTLAA------------------------------------------------ Hel308 Mzh (307) AFHHAGLNSEH------------RRMVEEGFRKNLIKMISSTPTLAA------------------------------------------------ Hel308 Nma (310) AFHHAGLSSTQ------------RSLVEDAFRDRLLKVISATPTLAA------------------------------------------------ Hel308 Nth (318) SSLELGIDIGA------------IDYVVQIDDTHTVSSLKQRLGRSG------------------------------------------------ Hel308 Pfu (298) AFHHAGLGRDE------------RVLVEENFRKGIIKAVVATPTLSA------------------------------------------------ Hel308 Sso (316) AYHHAGLSKAL------------RDLIEEGFRQRKIKVIVATPTLAA------------------------------------------------ Hel308 Tba (320) AFHHAGLGRAE------------RTLIEDAFREGLIKVLTATPTLAM------------------------------------------------ Hel308 Tga (299) AFHHAGLSRVE------------RTLIEDAFREGLIKVITATPTLSA------------------------------------------------ Hel308 Tsi (322) AFHHAGLGRTE------------RSIIEDAFREGLIKVITATPTLSA------------------------------------------------ Hel308 Mja (305) AFHHAGLTYQH------------RKIVEDAFRKRLIKVICCTPTLSA------------------------------------------------ Consensus (381) AFHHAGL R LVEDAFR LIKVI ATPTLAA 476 570 Hel308 Mbu (342) --------------------------------------GLNLPARRVIIRSYRRFDS-NFG--------------------------MQPIPVLE Hel308 Afu (335) --------------------------------------GVNLPARRVIVRSLYRFDG-YSK----------------------------RIKVSE Hel308 Csy (348) --------------------------------------GVNLPARRVVISSVMRYNS-SSGM-------------------------SEPISILE Hel308 Dth (448) NEAGKCIKSLEKDGELLASRDRSFYYTRARYPHKDVDLRGIGQTYNIFEHSTGEYLGEVDGVRAFKETHPGAVYLHMGETYVVQDLDLETFAVYA Hel308 Fac (325) --------------------------------------GVNLPARTVIIRDITRFSD---GY-------------------------SKPISGIE Hel308 Hla (350) --------------------------------------GVNTPARRVIVRDWRRYDG-EFGG-------------------------MKPLDVLE Hel308 Hpa (344) --------------------------------------GVNTPSRRVIVRDWRRYDG-DIGG-------------------------MQPLDVLE Hel308 Htu (345) --------------------------------------GVNTPARRVIVRDWRRFDP-SAGG-------------------------MAPLDVLE Hel308 Hvo (350) --------------------------------------GVNTPSRRVVVRDWQRYDG-DYGG-------------------------MKPLDVLE Hel308 Mac (342) --------------------------------------GLNLPARRVIIRSYRRYSS-DSG--------------------------MQPIPVLE Hel308 Mba (342) --------------------------------------GLNLPARRVIIRNYRRYSS-EDG--------------------------MQPIPVLE Hel308 Mbo (335) --------------------------------------GLNLPARRVIIRDYLRFSA-GEG--------------------------MQPIPVSE Hel308 Mev (342) --------------------------------------GLNLPARRVIVRNYKRYDP-NFG--------------------------MQPIPVLD Hel308 Mfe (342) --------------------------------------GLNLPCRRAIVKDLTRYT--NRG--------------------------MRYIPIME Hel308 Mfr (330) --------------------------------------GLNLPARRVIIRDYLRYSG-GEG--------------------------MRPIPVRE Hel308 Mhu (335) --------------------------------------GLNLPARRVIIRDYNRFAS-GLG--------------------------MVPIPVGE Hel308 Mig (343) --------------------------------------GINIPCRRAIVRDLMRFS--NGR--------------------------MKPIPIME Hel308 Min (324) --------------------------------------GLNLPCRRVIISELKRYT--RRG--------------------------LTYIPIME Hel308 Mma (339) --------------------------------------GLNLPCRRAIVRDIKRYS--QNG--------------------------LVDIPRME Hel308 Mmah (342) --------------------------------------GLNLPARRVIIRSYKRYDP-NAG--------------------------MQPIPVLD Hel308 Mmar (345) --------------------------------------GVNTPSRRVVVRDWRRYDG-SAGG-------------------------MAPLSVLE Hel308 Mmaz (342) --------------------------------------GLNLPARRVIIRSYRRYSS-DSG--------------------------MQPIPVLE Hel308 Mok (346) --------------------------------------GINMPCRRAIIRDLKRFS--SRG--------------------------YIPIPKME Hel308 Mth (337) --------------------------------------GLNLPARRVLIRSYKRYEA-GLG--------------------------TRPIPVME Hel308 Mzh (342) --------------------------------------GLNLPARRVIIRSYKRYDP-NFG--------------------------MKPIPVLE Hel308 Nma (345) --------------------------------------GVNTPARRVIVRDWRRFDP-SAGG-------------------------MAPLDVLE Hel308 Nth (353) --------------------------------------RKLGINQVLQVYSTINDSLVQSLA-------------------------VIDLLLEK Hel308 Pfu (333) --------------------------------------GINTPAFRVIIRDIWRYS--DFG--------------------------MERIPIIE Hel308 Sso (351) --------------------------------------GVNLPARTVIIGDIYRFNKKIAGY-------------------------YDEIPIME Hel308 Tba (355) --------------------------------------GVNLPSFRVIIRDTKRYS--TFG--------------------------WSDIPVLE Hel308 Tga (334) --------------------------------------GVNLPSFRVIIRDTKRYA--GFG--------------------------WTDIPVLE Hel308 Tsi (357) --------------------------------------GVNLPAYRVIIRDTKRYS--NFG--------------------------WVDIPVLE Hel308 Mja (340) --------------------------------------GLNLPCRRAIVKDLTRFIN---KG-------------------------MRYIPIME Consensus (476) GLNLPARRVIIRDYKRY G M PIPVLE 571 665 Hel308 Mbu (372) YKQMAGRAGRPHLDPYGESVLLAKTYDEF--AQLMENYVEADAEDIWSKLGTENALRTHVLSTIVNGFASTRQELFDFFGATFFAYQQ-DKWMLE Hel308 Afu (363) YKQMAGRAGRPGMDERGEAIIIVGKRDR---EIAVKRYIFGEPERITSKLGVETHLRFHSLSIICDGYAKTLEELEDFFADTFFFKQN--EISLS Hel308 Csy (379) YKQLCGRAGRPQYDKSGEAIVVGGVNAD----EIFDRYIGGEPEPIRSAMVDDRALRIHVLSLVTTSPGIKEDDVTEFFLGTLGGQQS-GESTVK Hel308 Dth (543) AKSEANYYTRPITEKYTEIVEVQATRATAAGELCLGRLKVTEHVSAYEKRLVRGQARIGLIPLDLPPLVFETQGMWFTLDSQVRRDVEDRRLHFM Hel308 Fac (354) IQQMIGRAGRPKYDKKGYGYIYAASPG---MLRVAEGYLTGELEPVISRMDSNSLIRFNVLALISSGIATDLKGIQDFYGKTLLAAQN-DIDGYE Hel308 Hla (381) VHQMCGRAGRPGLDPYGEAVLLANDADTK--EELFERYLWADPEPVRSKLAAEPALRTHVLATVASGFASTRDGLLSFLDNTLYATQTDDEGRLA Hel308 Hpa (375) VHQMFGRAGRPGLDPHGEAVLIAKSHDEL--QELFDQYVWADPEPVHSKLAAEPALRTHILATVASGFAGTEEELLDFLERTLYATQTDETGRLE Hel308 Htu (376) VHQMMGRAGRPGLDPYGEAVLLAKSHDES--EELFDRYIWADPEPVRSKLAAEPALRTHVLATIASGFARTRGGLLEFLEATLYASQSSEAGRLE Hel308 Hvo (381) VHQMMGRAGRPGLDPYGEAVLLAKDADAR--DELFERYIWADAEDVRSKLAAEPALRTHLLATVASGFAHTREGLLEFLDQTLYATQTDDPERLG Hel308 Mac (372) YKQMAGRAGRPRLDPYGEAVLLAKSYEEL--LFLFEKYIEAGAEDIWSKLGTENALRTHVLSTISNGFARTKEELMDFLEATFFAYQY-SNFGLS Hel308 Mba (372) YKQMAGRAGRPRLDPYGEAVLVAKSYKEF--VFLFENYIEANAEDIWSKLGTENALRTHVLSTISNGFARTYDELMDFLEATFFAFQY-SNFGLS Hel308 Mbo (365) YRQMAGRAGRPRLDPYGEAVLIAKEAEQV--PELFEVYIEAEAEDVHSRIAEPTALYTHVLSLVASGFAGTRGELTEFMNRSFYVHEHKQGRLIH Hel308 Mev (372) YKQMAGRAGRPSLDPYGESVLISHTYNEF--TDLLDRYIDAEPEDILSKLGTENALRTHVLSTIVNGFATTRQGMVDFMGSSFFAYQQ-QKWSLI Hel308 Mfe (371) IQQCIGRAGRLGLDPYGEGIIVAKNDR---DYLRSYQVLTQKPEPIYSKLSNQAVLRTQLLGLIATIEIRDEYDLEWFIRNTFYAYQYGNLREVA Hel308 Mfr (360) YRQMAGRAGRPHLDPYGEAILIAKTEYAV--NDLHEEYVEAPDEDVTSRCGEKGVLTAHILSLIATGYARSYDELMAFLEKTLYAYQHTGKKALT Hel308 Mhu (365) YHQMAGRAGRPHLDPYGEAVLLAKDAPSV--ERLFETFIDAEAERVDSQCVDDASLCAHILSLIATGFAHDQEALSSFMERTFYFFQHPKTRSLP Hel308 Mig (372) IHQCIGRAGRPGLDPYGEGIIFVKNER---DLERAEQYLEGKPEYIYSKLSNQAVLRTQLLGMIATREIENEFDLISFIKNTFYAHQYGNLGGVL Hel308 Min (353) VQQCIGRAGRPGLDEYGEGILVAKDER---DYLRALQCLTQKPEPIYSKLSNDSVLRTQILGLIATRYVLDEYDLEEFIKNTFYAYQYKNLDEIK Hel308 Mma (368) IQQCIGRAGRPGLDPYGEGIIYIKNER---DAEKAYEILTGSVENIYSKLANQKVLRIHILGLISTGEIKDGQNLVNFMKNTFYAHQFGNIGAVL Hel308 Mmah (372) YKQMAGRAGRPHLDPYGEAVVIVKTYEEF--TDVLERYISASAEDIWSKLGTENALRTHILSTIASGFANCHREILTFLGSTFFAHQQ-QSWNFE Hel308 Mmar (376) VHQMMGRAGRPGLDPYGEAVLIASSHDEV--DELFERYVWADPEPVRSKLAAEPALRTHILATVASGFARSRKGLLEFLEQTLYASQTDDSGQLE Hel308 Mmaz (372) YKQMAGRAGRPRLDPYGEAVLLAKSYEEF--VFLFEKYIEAGAEDIWSKLGTENALRTHILSTISNGFARTREELMDFLEATFFAFQY-SNFGLS Hel308 Mok (375) IHQCIGRAGRPNLDPYGEGIIYINNTENPELIENAKNYLIGNVEEIYSKLSNQKVLRTHMLGLITTGDIKNKNDLEEFIKNTFYAYQYQNTKKIL Hel308 Mth (367) YRQMAGRAGRPGLDPYGESLIMARSESEL--QKLMDHYVMGEPEDIWSKLASERALRTHVLATIASRFADSVDSLSRLMASTFYARQQ-DPSYLG Hel308 Mzh (372) YKQMAGRAGRPHLDPYGESVLIARSYDEF--MDIMENYVNADPEDIWSKLGTENALRTHVLSTIVNGFAYTYRGLMDFVKMTFFAYQK-EASDLH Hel308 Nma (376) VHQMMGRAGRPGLDPYGEAVLLAKSHDES--QELFDRYVWADPEPVRSKLAAEPALRTHVLATIASGFARTREGLLEFLEATLYASQSSEGGRLE Hel308 Nth (385) WIEPATEYPLPLDILFHQIISICHEANGVRLDPLIDNIKANAAFYKLKEEDINHVINYMIENDFLQLIRNSAELIVGLEGERLLRGKEFYAVFMT Hel308 Pfu (362) VHQMLGRAGRPKYDEVGEGIIVSTSDD---PREVMNHYIFGKPEKLFSQLSNESNLRSQVLALIATFGYSTVEEILKFISNTFYAYQRKDTYSLE Hel308 Sso (383) YKQMSGRAGRPGFDQIGESIVVVRDKEDV--DRVFKKYVLSDVEPIESKLGSERAFYTFLLGILSAEGNLSEKQLENFAYESLLAKQL-----VD Hel308 Tba (384) IQQMIGRAGRPKYDKEGEAIIVAKTEK---PEELMEKYIFGKPEKLFSMLSNDAAFRSQVLALITNFGVESFRELIGFLEKTFYYHQRKDLEILE Hel308 Tga (363) IQQMMGRAGRPRYDKYGEAIIVARTDE---PGKLMERYIRGKPEKLFSMLANEQAFRSQVLALITNFGIRSFPELVRFLERTFYAHQRKDLSSLE Hel308 Tsi (386) IQQMMGRAGRPKYDIEGQAIIIAKTEK---PEDLMKRYVLGKPEKLFSMLSNEASFRSQVLALITNFGVGNFKELVNFLERTFYYHQRKNLEALE Hel308 Mja (369) IQQCIGRAGRPGLDPYGEGIIVAKNDRDY---LRAYQALTQKPEPIYSKLSNQAVLRTQLLGLIATGEIRDEYDLEWFIRNTFYAHQYGNLREVA Consensus (571) I QM GRAGRP LDPYGEAVLIAKS D EL E YI ADPE IWSKLA E ALRTHVLALIASGFA T ELLDFL TFYAYQ L 666 760 Hel308 Mbu (464) EVINDCLEFLIDKAMVSET-E--------------------------------------DIEDASKLFLRGTRLGSLVSMLYIDPLSGSKIVDGF Hel308 Afu (453) YELERVVRQLENWGMVVEAAH-----------------------------------------------LAPTKLGSLVSRLYIDPLTGFIFHDVL Hel308 Csy (469) FSVAVALRFLQEEGMLGRR----------------------------------------------GGRLAATKMGRLVSRLYMDPMTAVTLRDAV Hel308 Dth (638) GGLHALEHGLIGCMPLIILTDRNDLGGIASPVHEQLHKG--------------AVFIYDGTPGGIGLCRQAFELGDRLVARAMGILSSCTCENGC Hel308 Fac (445) LAFESALYFLKDNDFITEEN----------------------------------------------DIYSATKFGRLTSDLYIDPVSSLILKKCL Hel308 Hla (474) AVTDTVLDYLAVNDFIERDRD------------------------------------------GGSESLTATGIGHTVSRLYLDPMSAAEMIDGL Hel308 Hpa (468) TVTQHVLDYLDRNGFLERDD-----------------------------------------------RLRATGLGHRVSQLYLDPMSAAEIIDGL Hel308 Htu (469) SVTDDVLDYLERNDFIERSR--DDEAEDSGEDDGPFTSAADLAEQ---------QAAK------REETLEATSLGHTVSRLYLDPMSAAEIVHGL Hel308 Hvo (474) QVTDRVLDYLEVNGFVEFEG----------------------------------------------ETIQATPVGHTVSRLYLDPMSAAEIIDGL Hel308 Mac (464) VVVDECLNFLRQEGMLEQDS----------------------------------------------DALISTMFGKLVSRLYIDPLSAALIAKGL Hel308 Mba (464) TVVNECLNFLRQEGMLEKD-----------------------------------------------DALIPTSFGKLVSRLYIDPLSAARIAKGL Hel308 Mbo (458) RAIDEALQFLITAEMVVEV----------------------------------------------GEHIGATELGTLVSRMYIDPRSAFAIVTTL Hel308 Mev (464) DVVDDCIEFLQDNEMIKD-----------------------------------------DG--ER---LYATRLGQVISTLYIDPLSGAIIIDKL Hel308 Mfe (463) KNINEVIRFLEEK--------------------------------------------------EFMIDFIPTELGKRVAELYIDPLSAKYMIDGL Hel308 Mfr (453) RTLDDALGFLTEAEMVTDL----------------------------------------------SGMLHATEYGDLTSRLYIDPHSAEIITTAL Hel308 Mhu (458) RLVADAIRFLTTAGMVEER----------------------------------------------ENTLSATRLGSLVSRLYLNPCTARLILDSL Hel308 Mig (464) RNIKEVINFLEEN--------------------------------------------------DFIADYFPTKLGKRVSELYIDPLSAKIIIDGL Hel308 Min (445) KKIKEIIEFLEDCN--------------------------------------------------FIKNFEVTPLGKKVSNLYLDPLSAKIMIDNI Hel308 Mma (460) LNVSEVVEFLEKNKFLETTIHKKTENKVRELSFDS------S-NN---LVLDSKETSFDLTNPNSNIEFRSTKLGKRISELYIDPMSSEIIIEEL Hel308 Mmah (464) ELLEDCLIFLKNEGMLEQD-N--------------------------------------ET-------IRATELGKMISSLYIDPLSASKIIRGL Hel308 Mmar (469) RVVDDVLTYLQRNDFLEIEAG----------------------------------------------ELDATSLGHTVSRLYLDPMSAAEIVDGL Hel308 Mmaz (464) AVVDECLDFLRREGMLEKDP----------------------------------------------DALVSTVFGKLVSRLYIDPLSAALIAKGL Hel308 Mok (470) ENIYEITNFLEKNGFIELNYRRDENKDKSNNSHNNKKNISNTNNSIKMLVLDNNNSLTIKSRHEEDVYYNITPLGKKVSELYIDPLSAEYIIDGL Hel308 Mth (459) ETIASVLEFLVRSDMIDKD-------------------------------------------------LTPTPLGALVSRLYIDPLSAMVMIQEI Hel308 Mzh (464) DVIEECVRFLIDNEMIISD-S--------------------------------------NDILPES-AFRSTATGKLISMLYIDPLSGSLIMDGI Hel308 Nma (469) RVTDDVLSYLERNDFIERSGGPEDTLNSEADAASAFTSAADLADS---------DGGDSGGTTGQEEDLEATSLGHTVSRLYLDPMSAAEIVHGL Hel308 Nth (480) QEEFEVREGIRKIGSIDKS-------------------------------------------LMVSEGDNIILAGQLWTIKNIDIERDIIYVAKA Hel308 Pfu (454) EKIRNILYFLLEN---------------------------------------------EFIEISLEDKIRPLSLGIRTAKLYIDPYTAKMFKDKM Hel308 Sso (471) VYFDRAIRWLLEHSFIKEE----------------------------------------------GNTFALTNFGKRVADLYINPFTADIIRKGL Hel308 Tba (476) GKAKSIVYFLLEN---------------------------------------------EFIDIDLNDSFIALPFGIRTSQLYLDPLTAKKFKDAL Hel308 Tga (455) YKAKEVVYFLIEN---------------------------------------------EFIDLDLEDRFIPLPFGKRTSQLYIDPLTAKKFKDAF Hel308 Tsi (478) GKAKSIVYFLFEN---------------------------------------------EFIDIDLNDQFMPLPLGIRTSQLYLDPVTAKKFKDAF Hel308 Mja (461) KNINEVIRFLEENEFI--------------------------------------------------IDFMPTELGKRVSELYIDPLSAKFIIDGL Consensus (666) I EVL FL N I L AT LG VS LYIDPLSA IIDGL 761 855 Hel308 Mbu (520) KDIGKSTGGNMGSLEDDKG----------------------------------------------------DDITVTDMTLLHLVCSTPDMRQLY Hel308 Afu (501) SRMELS-----------------------------------------------------------------------DIGALHLICRTPDMERLT Hel308 Csy (518) GEASPGR--------------------------------------------------------------------MHTLGFLHLVSECSEFMPRF Hel308 Dth (719) PGCIHSPKCGSGNR------------------------------------------------------------PLDKEAAMHMLAVLAGERCGE Hel308 Fac (494) DLEFS------------------------------------------------------------------------EELYLYYISKTPDMLTFN Hel308 Hla (527) RSVARDAADTGASAEADNG-EFVRTGDADDASGGDEPGFGTYTRAGDDESGER------ETENEETDEEETEASEVTPLGLYHLISRTPDMYELY Hel308 Hpa (516) RDADG---------------------------------------------------------------------KPTALGLYHLVSRTPDMYQLY Hel308 Htu (547) ERADER---------------------------------------------------------------------PTALGLYQLVSRTPDMYELY Hel308 Hvo (523) EWAADHRTEKLRALAGETPEKPTRDRSESDESGGFQRASEMVADDGDGGGGEDGVGANGDGDSDDADGVETDRTYPTPLGLYHLVCRTPDMYQLY Hel308 Mac (513) REAGT----------------------------------------------------------------------LTELTLLHLVCSTPDMRLMY Hel308 Mba (512) KGAKS----------------------------------------------------------------------LSELTLLHLVCSTPDMRLLY Hel308 Mbo (507) REQEK----------------------------------------------------------------------YADLGLIQLICTTPDMPTLY Hel308 Mev (513) KKADK----------------------------------------------------------------------VTDMTMLHIICSTPDMRQLY Hel308 Mfe (508) NEMENED----------------------------------------------------------------------DIYYLYLISKTLEMMPNL Hel308 Mfr (502) REEGE----------------------------------------------------------------------LTDLALLQLLCMTPDMFTLY Hel308 Mhu (507) KSCKT----------------------------------------------------------------------PTLIGLLHVICVSPDMQRLY Hel308 Mig (509) KEMGNVDNE--------------------------------------------------------------------ELYYLYLISKTLEMMPLL Hel308 Min (490) EVKDDLH-------------------------------------------------------------------------LLYILCKCIEMKPLL Hel308 Mma (545) HELKKKCDQLDR--------------------------------------------------------------SKIDQYLFYLISKTNEMRPLL Hel308 Mmah (513) EKTTH----------------------------------------------------------------------VTDMTLLQLICSTPDMRLLY Hel308 Mmar (518) RDWERGASDSTSASGSPAD----AQAEP-PANSGFTTASELAEDADESDADRD-------------------PDDISALGLYHLVSRTPDMYQLY Hel308 Mmaz (513) REAGT----------------------------------------------------------------------LTELTLLHLICSTPDMRLMY Hel308 Mok (565) KNLHKKTLSNPKNM------------------------------------------------------------ECYILHILYIISKTTEMQPVL Hel308 Mth (505) RGIRR----------------------------------------------------------------------PTVLTLLHVITMTPDMELLF Hel308 Mzh (519) RKADY----------------------------------------------------------------------FEDITMMHLICSTPDMKNLY Hel308 Nma (555) EDADER---------------------------------------------------------------------PTALGLYQLVSRTPDMYELY Hel308 Nth (532) VDGKPPK--------------------------------------------------------------------YSGGGFILNPKIPERMHKIL Hel308 Pfu (504) EEVVKDPN---------------------------------------------------------------------PIGIFHLISLTPDITPFN Hel308 Sso (520) EGHKAS----------------------------------------------------------------------CELAYLHLLAFTPDGPLVS Hel308 Tba (526) PQIEENPN---------------------------------------------------------------------PLGIFQLLASTPDMGTLS Hel308 Tga (505) PAIERNPN---------------------------------------------------------------------PFGIFQLIASTPDMATLT Hel308 Tsi (528) EKLEKNPN---------------------------------------------------------------------PLGIFQLLASTPDMSSLR Hel308 Mja (506) EEMENEE----------------------------------------------------------------------EIYYLYLISKTLEMMPNL Consensus (761) LGLLHLIS TPDM LY 856 950 Hel308 Mbu (563) LRNTDYTIVNEYIVAHSDEFH---EIPDKLKETDYEWFMGEVKTAMLLEEW---------------VTEVSAEDITRHFNVGEGDIHALADTSEW Hel308 Afu (525) VRKTDSWVEEEAFRLRKELSY----YPSDFS-VEYDWFLSEVKTALCLKDW---------------IEEKDEDEICAKYGIAPGDLRRIVETAEW Hel308 Csy (545) ALRQKDHEVAEMMLEAGRGELLR---P--------VYSYECGRGLLALHRW---------------IGESPEAKLAEDLKFESGDVHRMVESSGW Hel308 Dth (754) AKRKDVSCRIETDEGSMEIDSG-YTKSDQAELPYAVLDIETRYSAQEVGGWGNCHRMGVSFAVVFDSRNQEFVTFDQEQAADLGSFLEDFSLVVG Hel308 Fac (517) YRASDYEYLEEFLDRHNISDFS--EESMGAAKTAIILNEW--------------------------INEVPINTIAETFGIGPGDIQAKASSADW Hel308 Hla (615) LKSGDRETYTELCYERETEFLG--DVPSEYEDVRFEDWLASLKTARLLEDW---------------VNEVDEDRITERYGVGPGDIRGKVDTAEW Hel308 Hpa (542) LRSGDRERYTEIAYEREPEFLG--HMPSEFEDNAFEDWLSALKTARLLEDW---------------ASELDEDRITERYAIGPGDIRGKVETAQW Hel308 Htu (573) LRSGEDEKFGELFYERETELLG--DAPSEYEEDRFEDWLAALKTGKLLEDW---------------ADETDEETITDRYKIGPGDLRGKVDTAEW Hel308 Hvo (618) LKSGDRETYTELCYEREPEFLG--RVPSEYEDVAFEDWLSALKTAKLLEDW---------------VGEVDEDRITERYGVGPGDIRGKVETSEW Hel308 Mac (538) MRSQDYQDINDFVMAHAEEFS---KVPSPFNIVEYEWFLSEVKTSLLLMDW---------------IHEKPENEICLKFGTGEGDIHTTADIAEW Hel308 Mba (537) MRSHDYQDINDYVMAHASEFV---KVPSPFDTTEYEWFLGEVKTSLLLLDW---------------IHEKSENEICLKFGTGEGDIHSIADIAEW Hel308 Mbo (532) AKNADLPALSRMLEVRGADIW---LPP-PLDDDAAETYYRAVKTAMLLSDW---------------TDELSEEKICERYGVGPGDVFGMVENINW Hel308 Mev (538) LRSKEYEKINEYVMTHSDEFV---EVPNPFKSIEYEWFLGEVKTALLINEW---------------IDEKTLDDITAEFGVGEGDINALSDISEW Hel308 Mfe (533) RVYKSEE--LNLIDEMENLG------IKSFE----IEDLEAFKTAKMLYDW---------------ISEVPEDEILKKYKIEPGILRYKVENAVW Hel308 Mfr (527) VKKNDLGTLEKFFFEHEEEFR---T---EFSYDEMEDFFRSLKTAMLLSDW---------------TDEIGDDTICTRFGVGPGDIFNAVQGISW Hel308 Mhu (532) LKAADTQLLRTFLFKHKDDLI---LPL-PFEQEEEELWLSGLKTALVLTDW---------------ADEFSEGMIEERYGIGAGDLYNIVDSGKW Hel308 Mig (536) RVNSFEE--LDLILEMEEAG------IYDRT----YDDLAAFKNAKMLYDW---------------INEVPEDEILKKYKIEPGILRYKVEQAKW Hel308 Min (512) RVYRKEE--EELAEELLNYE------I-FIS----YENLEEFKTAKMLYDW---------------INEVPEDEILKTYKVEPGILRYKVEVAKW Hel308 Mma (578) RIRPNEE--LDLILEMDKMG------LKDYS----IENIEAFKNSKMFCDW---------------VSEIPEEIILEKYGVEPGILRYKVEQAKW Hel308 Mmah (538) LRNRDYEIINDYVMNHTEEFI---EVPSPFKQIEYEWFLSEVKTALLLLEW---------------INEKSLEKIVENYQVGEGDIYASSDIAEW Hel308 Mmar (589) LRSGDREEYEMELFEREEELLG--PTPSEFEEGRFEDWLSALKTARLLEDW---------------ATEVDEATITDRYGVGPGDIRGKVETAQW Hel308 Mmaz (538) MRSQDYQEVNDYVMAHAGEFS---KVPNPFNIAEYEWFLGEVKTSLLLMDW---------------IHEKPENEICLKFGIGEGDIHATADIAEW Hel308 Mok (600) RVRRKEE--NDLINDMIKLDIDVDDVIYGIS----SENLEYFKNAKLFYDW---------------INEIPEEELLLGYNIEPGILRYNVEQAKW Hel308 Mth (530) VQQS-DNWLEDFISEHSSELG---NEKN------FDWLLREVKTASMLMDW---------------INEVHEDRIEDRYSISPGDLVRIAETAEW Hel308 Mzh (544) MRSSDYENVNMYVLQNKDKFI---SMPSPFKMIEYEWFLGEVKTALLLLDW---------------INEVPADDICKKYGIGEGDIRMFSETAVW Hel308 Nma (581) LRSGEDEKFGELYYERERELLG--DAPSEFEEERFEDWLAALKTGKLLEDW---------------ATEDDEEQITERYKIGPGDLRGKVDTAEW Hel308 Nth (559) CERKNFEFIDNMAQNHLEEQR---------------------KPFELYNIK---------------PNERVIWNNGDEILFETYTGTKIFQTLAW Hel308 Pfu (530) YSKREFERLEEEYYEFKDRLYFDDPYISGYDPYLERKFFRAFKTALVLLAW---------------INEVPEGEIVEKYSVEPGDIYRIVETAEW Hel308 Sso (545) VGRNEEEELIELLEDLDCELL----IEEPYEEDEYSLYINALKVALIMKDW---------------MDEVDEDTILSKYNIGSGDLRNMVETMDW Hel308 Tba (552) IKRKEQESYLDYAYEMEDYLYRSIPYWEDYE---FQKFLSEVKTAKLLLDW---------------INEVSEAKLIEAYGIDTGDLYRIIELADW Hel308 Tga (531) ARRREMEDYLDLAYELEDKLYASIPYYEDSR---FQGFLGQVKTAKVLLDW---------------INEVPEARIYETYSIDPGDLYRLLELADW Hel308 Tsi (554) VKRKEQEDLLDYAYEMEEYLYQNIPYWEDYK---FEKFLGETKTAKLLLDW---------------INEVNDVKILETYEIDTGDLYRILELVDW Hel308 Mja (531) RVYNSEE--LNLIDEMDSLGIK----------SFEIEDLEAFKTAKMLYDW---------------INEVPEDEILKRYKIEPGILRYKVENAVW Consensus (856) LR D E L E I E E F FE FL VKTA LL DW I EV ED I ERYGIGPGDL VE AEW 951 1045 Hel308 Mbu (640) LMHAAAKLAELLGVEYSS--------HAYSLEKRIRYGSGLDLMELVGIRGVGRVRARKLYNAGFVS---------VAKLKGADISVLSKLVGP- Hel308 Afu (600) LSNAMNRIAEEVG-N--T--------SVSGLTERIKHGVKEELLELVRIRHIGRVRARKLYNAGIRN---------AEDIVRHREKVASLIGRG- Hel308 Csy (614) LLRCIWEISKHQERPDLLG-------ELDVLRSRVAYGIKAELVPLVSIKGIGRVRSRRLFRGGIKG---------PGDLAAVPVERLSRVEGIG Hel308 Dth (848) FNLLKFDYRVLQGLSDYDFSSLPTLDMLREIEARLGHRLSLDHLARHTLGTNKSANGLMALKWWKEGELDKIVEYCRQDVSVTRDLYLFGRDKGY Hel308 Fac (584) ISYSLYRLGSMFDKENEN--------NLLHLNIRIKEGVKEEIIRIIEIPQVGRVRGRRLYNNGFKS---------IDDIANARVEDISRIFGFS Hel308 Hla (693) LLRAAETLARDVEGVDGDVVV-----AVREARKRIEYGVREELLDLAGVRNVGRKRARRLFEAGIET---------RADLREADKAVVLGALRGR Hel308 Hpa (620) LLNAAERLAAELQRDDAEGIPSATTTAVREARKRVEYGVEEELLDLAGVRNVGRKRARRLYEAGIES---------RADLREADKSVVLGALRGR Hel308 Htu (651) LLGAAESLAAEIDSEWTV--------AVREARARVEHGVGEELLELVSVGGVGRKRARRLYDAGIEE---------PADLRSADKGIVLSVLKG- Hel308 Hvo (696) LLGAAERLATELD---LDSVY-----AVREAKKRVEYGVREELLDLAGVRGVGRKRARRLFEAGVET---------RADLREADKPRVLAALRGR Hel308 Mac (615) IMHVATQLARLLDLKGAK--------EAAELEKRIHYGAGPELMDLLDIRGIGRVRARKLYGAGFKS---------TADLAGATPEKVAALVGP- Hel308 Mba (614) IMHVTSQLAGLLDLKGAR--------EAAELEKRIHYGAAPELIDLLNIRGIGRVRARKLYEAGFKS---------SAELAEVDPEKVAALLGP- Hel308 Mbo (608) LLHATSQLARMFVPKFYG--------QIADCEICMKNGIRRELLPLVRLRGIGRVRARRLFNNGITS---------PEELSRHKKEDLVKILGS- Hel308 Mev (615) LMHSAVNLANLTDLDAD---------KAQELEKRIHHGVNKDLIQLVSISNIGRVRARKLYEAGIQS---------VSDIKNTKLHILSNYLGR- Hel308 Mfe (601) LMHALKEMAKIIGKN---------SEIPEKLEIRLEYGAKEDIIELLNVKYIGRVRARKLYNAGIRN---------VEDIINNPSK---VASIIG Hel308 Mfr (601) LLHASGRLARLVAPEHRD--------AVEETTLRVRHGIRRELIPLVRVKGIGRVRARRLFNNGITG---------PELLAAADPSVVGHIVGG- Hel308 Mhu (608) LLHGTERLVSVEMPEMSQ--------VVKTLSVRVHHGVKSELLPLVALRNIGRVRARTLYNAGYPD---------PEAVARAGLSTIARIIGE- Hel308 Mig (604) MIYSTKEIAKLLNRN---------IDTLSKLEIRLEYGAKEDIIELLKIKYVGRARARKLYDAGIRS---------VEDIINNPKK---VASLLG Hel308 Min (579) LSYSLKEIAKILNKEVP------------NLELRLEYGAKEELLELLKIKYIGRVRARKLYSAGIRN---------REDIIKNPKK---VANILG Hel308 Mma (646) MIYSTKEIAKLIHLDNSE-----IYKSLLKMEVRIEYGAKEELIELLNVKNVGRIRSRKLYDAGIRS---------KIEINKNPEK---ILELFG Hel308 Mmah (615) LMHATQRIASRINPQLET--------ECAKLEKRIHYGAGSELIELVEIPNVGRARARKLFKKGYRS---------RQKLATADEKQLAGIVGP- Hel308 Mmar (667) LLGAAESLASEVDLDAAR--------AISEARIRVEHGVREELVDLAGVRGVGRKRARRLFQAGITD---------RAQLRDADKAVVLAALRGR Hel308 Mmaz (615) IMHVTAQLAGLLDLKGAK--------EASELEKRIRYGAAPELMDLLDIRSVGRVRARKLYEAGFKS---------TAELAAASPEHIAVLVGP- Hel308 Mok (674) MIHSAKEIFNLLNIDNKV-----IKDCLNDLEIRMEYGAKQDIIELLKIKHIGRARARILYNAGIKN---------ANDIINNQKN---IINLLG Hel308 Mth (600) LMSALHRISKHMDLGVTY--------LAERLALRIHYGAGDELLQLLELKGIGRVRARKLYQAGYRS---------LEDLKAADKSTLSEILGP- Hel308 Mzh (621) LMHATSRLSGLLKVSEASE-------KSKELEKRLSYGINSELVNIVALKGIGRVRARKIYENGYRS---------IDDLKKADPLKLSKIVGS- Hel308 Nma (659) LLGAAESLASEIDSEWAV--------AVREARARVEHGVGEELLELVSVSGIGRKRARRLYAAGIEE---------PAALRSADKGVILHVLKG- Hel308 Nth (618) ILRSYNVNIKEIDGIGRIN-----IEGGIDLPGVLQDIKETDWRPEYLLDFTLEQEKFKSKFSPYLP---------KDLQDKMHIAHLVDIEGVK Hel308 Pfu (610) LVYSLKEIAKVLG-AYE------IVDYLETLRVRVKYGIREELIPLMQLPLVGRRRARALYNSGFRS---------IEDISQARPEELLKIEGIG Hel308 Sso (621) LTYSAYHLSRELKLNEHAD-------KLRILNLRVRDGIKEELLELVQISGVGRKRARLLYNNGIKE---------LGDVVMNPDKVKNLLGQK- Hel308 Tba (629) LMYSLIELAKVLNAGGE------TIKYLRRLHLRLKHGVREELLELVELPMIGRRRARALYNAGFKN---------VNDIVKAKPSELLAVEGIG Hel308 Tga (608) LMYSLIELYKLFEPKEE------ILNYLRDLHLRLRHGVREELLELVRLPNIGRKRARALYNAGFRS---------VEAIANAKPAELLAVEGIG Hel308 Tsi (631) LMYSLIELYKLFDPKPE------VLDFLKKLHIRVKHGVREELLELITLPMIGRKRARALYNAGFKG---------IDDIVRAKASELLKVEGIG Hel308 Mja (599) IMHALKEIAKLIGKSSDI---------PEKLEIRLEYGAKEDIIELLSIKYIGRVRARKLYNAGIRS---------IEDIINNPSK---VASIIG Consensus (951) LMHA LAKLL L EL IRI YGVKEELLELV IR IGRVRARKLY AGIRS DL A L ILG 1046 1140 Hel308 Mbu (717) -KVAYNILSGIGVRVNDKHFNSAPISSNTL--------------------TLLDKNQKTFNDFQ------------------------------- Hel308 Afu (674) --IAERVVEGISVKSLNPESAAALEHHHHHH --------------------------------------------------------------- Hel308 Csy (693) ATLANNIKSQLRKGG ------------------------------------------------------------------------------- Hel308 Dth (943) LLFKNKAGKKVRIPVSWQDTAFQV----------------------------------------------------------------------- Hel308 Fac (662) TKLAKDIIENAGKLNNRYYR--------------------------------------------------------------------------- Hel308 Hla (774) ERTAERILEHAGREDPSMDDVRPDKSASAAATAGS-------------ASDEDGEGQASLGDFR------------------------------- Hel308 Hpa (706) KKTAENILENVGRQDPSLDDVEADAET---------AA--------TSARATNDGGQQSLGDFE------------------------------- Hel308 Htu (728) EKTAENILENAGREDPSMDGVEPADGGPAVGAATNGSSGGSETDETGRADAAESDDSQSSLGDF------------------------------- Hel308 Hvo (774) RKTAENILEAAGRKDPSMDAVDEDDAPDDAVPDDA---G--------FETAKERADQQASLGDFEGS---------------------------- Hel308 Mac (692) -KIAERIFRQIGRREAVSEISDSERLEKS------------------------SQDGQSTISDF------------------------------- Hel308 Mba (691) -KIADRIFKQIRGRGTSSGIIASEPPEKS------------------------PYSGQKTISDY------------------------------- Hel308 Mbo (685) -GIAEQVLEQLHPSKDTGKKEPPSGDKNTN------------------------PG-QSTLFHFG------------------------------ Hel308 Mev (691) -KTAYKVLEQLGVEPEDNQQIDEEPESIKSY-------------------SGNDQGQKTFNDF-------------------------------- Hel308 Mfe (675) EKITKKILEDLG--IKFGQ--------------------------------------QKLIF--------------------------------- Hel308 Mfr (678) -KTAESII -------------------------------------------------------------------------------------- Hel308 Mhu (685) -GIARQVIDEITGVKRSGIHSSDDDYQQKT------------------------PE-LLTDIPGIGKKMAEKLQNAGIITVSDLLTADEVLLSDV Hel308 Mig (678) EKIAKKILGELG--MKFGQ--------------------------------------QTLQI--------------------------------- Hel308 Min (650) EKISKKIFEELG--VRYGQ--------------------------------------QRLI---------------------------------- Hel308 Mma (724) EKIGKKILGEHG--MKYGQ--------------------------------------QTLLNFN------------------------------- Hel308 Mmah (692) -KIAQKILSYLGRETDSNGYVEPETLENK-------------------------KQ-QKTFQDFI------------------------------ Hel308 Mmar (745) RKTAENVLENAGHRDPSMEGVEPAPDVSVDLNDGADGD---------ASAESTANDDQASLGDF------------------------------- Hel308 Mmaz (692) -KITERIFKQIGRREAVSEFSDIEPLEKG------------------------SSDGQRTISDY------------------------------- Hel308 Mok (752) EKIARKILSELGVDTKFGQ---------------------------------------MRLSI-------------------------------- Hel308 Mth (677) -KIAEGVISQLK-EPGVSA---------------------------------------------------------------------------- Hel308 Mzh (699) -KISQKILKQLDIDVDISEIKEKDSDTVP-E--------------------P--ESSQKTISDFT------------------------------ Hel308 Nma (736) EKTAENILENAGREEPSMDGVEPIPVEGGSGSGSSNSSGSSEPNADANATEDDADDNQSSLGDF------------------------------- Hel308 Nth (699) TFLENKKIKEIKL --------------------------------------------------------------------------------- Hel308 Pfu (689) VKTVEAIFKFLGKNVKISE-----------------------------------KPRKSTLDYFLKS---------------------------- Hel308 Sso (699) -LGEKVVQEAARLLNRFH----------------------------------------------------------------------------- Hel308 Tba (709) VKVLERIYRHFGVELPLLKNIKDPDKPEDKPKEKP-------------------KPKKGTLDYFLK----------------------------- Hel308 Tga (688) AKILDGIYRHLGIEKRVTE-----EK----------------------------PKRKGTLEDFLR----------------------------- Hel308 Tsi (711) IGVIEKIYQHFGVELPTNE-----KK--------K-------------------KVKKGTLDEFFK----------------------------- Hel308 Mja (673) EKIAKKILDELGVKFGQQKLSFSGGSAWSHPQFEKGGGSGGGSGGSAWSHPQFEK----KL---------------------------------- Consensus (1046) KIAEKIL LG TL F 1141 1186 Hel308 Mbu (761) ---------------------------------------------- Hel308 Afu (703) ---------------------------------------------- Hel308 Csy (708) ---------------------------------------------- Hel308 Dth (967) ---------------------------------------------- Hel308 Fac (682) ---------------------------------------------- Hel308 Hla (825) ---------------------------------------------- Hel308 Hpa (753) ---------------------------------------------- Hel308 Htu (792) ---------------------------------------------- Hel308 Hvo (830) ---------------------------------------------- Hel308 Mac (731) ---------------------------------------------- Hel308 Mba (730) ---------------------------------------------- Hel308 Mbo (724) ---------------------------------------------- Hel308 Mev (734) ---------------------------------------------- Hel308 Mfe (697) ---------------------------------------------- Hel308 Mfr (685) ---------------------------------------------- Hel308 Mhu (754) LGAARARKVLAFLSNSEKENSSSDKTEEIPDTQKIRGQSSWEDFGC Hel308 Mig (700) ---------------------------------------------- Hel308 Min (671) ---------------------------------------------- Hel308 Mma (748) ---------------------------------------------- Hel308 Mmah (730) ---------------------------------------------- Hel308 Mmar (800) ---------------------------------------------- Hel308 Mmaz (731) ---------------------------------------------- Hel308 Mok (776) ---------------------------------------------- Hel308 Mth (694) ---------------------------------------------- Hel308 Mzh (740) ---------------------------------------------- Hel308 Nma (800) ---------------------------------------------- Hel308 Nth (712) ---------------------------------------------- Hel308 Pfu (721) ---------------------------------------------- Hel308 Sso (716) ---------------------------------------------- Hel308 Tba (756) ---------------------------------------------- Hel308 Tga (721) ---------------------------------------------- Hel308 Tsi (745) ---------------------------------------------- Hel308 Mja (730) ---------------------------------------------- Consensus (1141) ----------------------------------------------