RECOMBINANT STRAIN HAVING MODIFIED SUGAR METABOLIC PATHWAY AND METHOD FOR SCREENING SUGAR ISOMERASE USING SAME
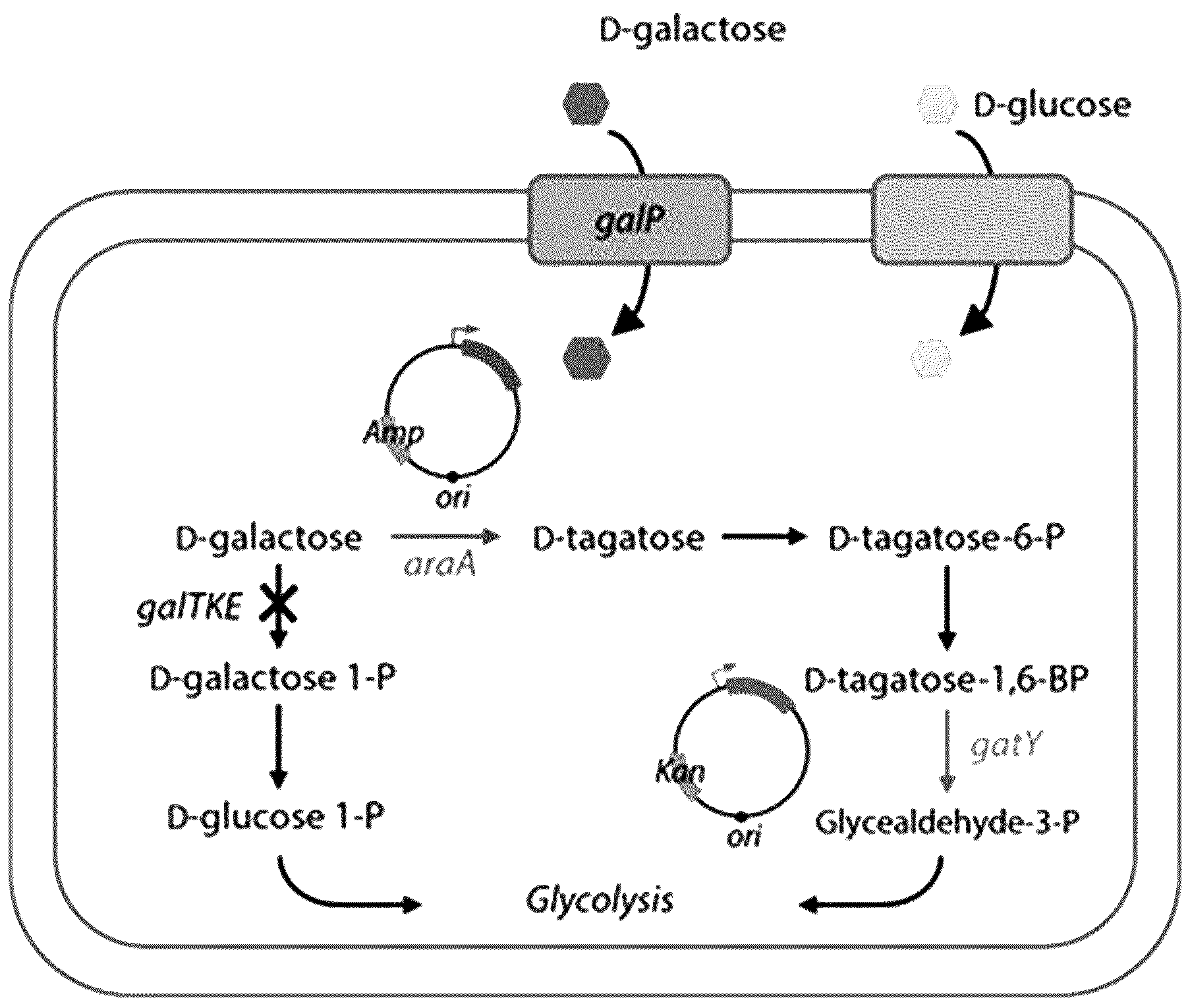
본 발명은 균주에 타 균주 유래의 효소를 도입하여 변형된 당 대사 경로를 갖는 재조합 균주에 관한 것이며, 이를 이용하여 유용물질을 얻거나, 유용물질을 생산할 수 있는 다양한 변이체 및 변이 효소를 초고속 스크리닝 할 수 있는 방법에 관한 것이다. L-아라비노오스 이성화효소(L-arabinose isomerase; EC 5.3.1.5)는 생체내( D-타가토오스는 D-갈락토오스의 이성질체이며 과일, 우유, 치즈 등에 존재하는 천연 당류이다. D-타가토오스는 다양한 건강 기능적 특성과 설탕과 매우 유사한 단맛을 가지고 있기 때문에 여러 제품 적용 시 건강과 맛을 동시에 만족시킬 수 있는 대체 감미료로 사용된다. 한편, 일반적으로 효소의 활성 및 구조적 안정성을 증진시키거나 새로운 기질에 대한 활성을 부여하는 등 원하는 목적에 부합하도록 효소의 특성을 변환시키는 개량기술로 분자진화 (directed evolution) 기술이 사용되고 있다. 이러한 기술을 수행하기 위한 변이주 라이브러리를 제조하기 위해 가장 일반적인 방법으로 많이 사용되는 것은 error-prone PCR 방법으로 PCR 수행시 DNA 중합효소의 에러발생율을 조절하여 무작위적으로 돌연변이를 도입하는 방법이다. 이렇게 만들어진 변이주들을 이용하여 단백질을 발현시킨 후, 활성이 좋은 변이주를 선별함으로써 우수한 활성을 갖는 개량 효소를 수득하게 되는데 원하는 효소의 특징과 목적에 맞는 효율적인 스크리닝 기술을 개발하는 것이 분자진화의 핵심기술이라 할 수 있다. 효소개량 라이브러리 구축을 위하여 무작위 돌연변이법 및 포인트 돌연변이법 등 다양한 변이주 제작을 위한 시도가 있었으나, 이러한 방법은 개량된 효소의 초고속 탐색방법의 부재로 107 이상의 변이체를 스크리닝하는 분자진화기술을 적용하기엔 한계가 있다. 즉, 당전환 효소의 단백질 공학기술 적용을 위해서 초고속 탐색이 불가한 기존의 방법을 대체할 수 있는 신규한 효소 탐색법 연구가 필요한 실정이다. 본 발명자들은 유용물질을 생산하기 위한 새로운 대사경로 구축과, 새로운 당전환 효소의 빠른 스크리닝법에 관한 연구를 수행하던 중, D-타가토오스-1,6-비스포스페이트 알돌라아제(D-tagatose 1, 6-bisphosphate aldolase)를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자를 D-갈락토오스 비대사성 균주에 도입하는 경우, 새로운 당 대사 경로를 갖는 균주를 수득할 수 있으며, L-아라비노오스 이성화효소를 코딩하는 유전자에 무작위 변위를 유발한 라이브러리 구축과 이를 이용한 균주 성장 측정을 통해 신규한 당 이성화 효소를 빠르게 스크리닝 할 수 있음을 확인하고 본 발명을 완성하였다. 따라서 본 발명의 목적은 새롭게 구축된 당 대사 경로를 이용하여 새로운 당 이성화능 및 당 대사능을 갖는 균주를 신속하게 스크리닝 하고, 이를 통해 신규한 변이 당 이성화 효소를 초고속으로 스크리닝 하는 방법을 제공하는 것이다. 상기와 같은 목적을 달성하기 위하여 본 발명은 1) 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계; 2) 제1당 대사능 및 제2당 이성화능이 결여된 균주에 상기 1) 단계에서 선별된 효소를 암호화하는 유전자를 도입하여 제1당 대사성 재조합 균주를 제조하는 단계; 3) 제2당 이성화 효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계; 4) 상기 3) 단계에서 수득된 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계에서 제조된 제1당 대사성 재조합 균주에 도입하여 균주 라이브러리를 얻는 단계; 및 5) 제2당을 유일 탄소원으로 포함하는 제한 배지에서 상기 4) 단계에서 수득된 라이브러리의 균주를 배양하고 균의 성장 및 생장속도를 확인하는 단계; 를 포함하며, 상기 제1당과 제2당은 상호 이성질체인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 및 제1당 대사능을 갖는 변이체의 초고속 스크리닝 방법을 제공한다. 또한 본 발명은 1) 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계; 2) 제1당 대사능 및 제2당 이성화능이 결여된 균주에 상기 1) 단계에서 선별된 효소를 암호화하는 유전자를 도입하여 제1당 대사성 재조합 균주를 제조하는 단계; 3) 제2당 이성화 효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계; 4) 상기 3) 단계에서 수득된 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계에서 제조된 제1당 대사성 재조합 균주에 도입하여 균주 라이브러리를 얻는 단계; 5) 제2당을 유일 탄소원으로 포함하는 제한 배지에서 상기 4) 단계에서 수득된 라이브러리의 균주를 배양하고 높은 성장능을 갖는 균주를 선별하는 단계; 및 6) 상기 5) 단계에서 선별된 균주에 도입된 변이 효소를 확인하는 단계; 를 포함하며, 상기 제1당과 제2당은 상호 이성질체인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 을 갖는 변이 당이성화 효소의 초고속 스크리닝 방법을 제공한다. 또한 본 발명은 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자를 포함하는, 재조합 벡터를 제공한다. 또한 본 발명은 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자가 도입된, D-갈락토오스 대사능을 갖는 재조합 균주를 제공한다. 또한 본 발명은 D-갈락토오스 비대사성 균주에 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자를 도입하는 단계; 를 포함하는 D-갈락토오스 대사능을 갖는 재조합 균주의 제조방법을 제공한다. 또한 본 발명은 서열번호 2 내지 4로 이루어진 군에서 선택된 1종의 아미노산 서열로 표시되는, 당 이성화능을 갖는 L-아라비노오스 이성화효소 변이효소를 제공한다. 본 발명에 따른 초고속 스크리닝 방법, 재조합 벡터 및 균주를 이용하면, 새로운 당 대사경로를 균주에 구축하여 D-갈락토오스로부터 D-타가토오스를 효과적으로 수득할 수 있을 뿐만 아니라, 무작위적으로 변형된 당 이성화 효소를 도입한 후, 세포 성장 기반 초고속 탐색법을 수행함으로써, 유용한 당이성화 변이효소를 빠르게 스크리닝할 수 있는 장점이 있다. 도 1은 본 발명의 “세포성장 기반 초고속 탐색법” (cell growth-associated screening system)을 구축하기 위하여 균주에 도입되는 새로운 당 대사경로를 나타낸 모식도이다. 도 2는 복합 (LB) 배지와 포도당(glc) 및 D-갈락토오스(gal) 가 유일 탄소원으로 첨가된 제한 (M9) 배지에서의 대장균 균체 성장곡선을 나타낸 도이다. 도 3은 D-타가토오스 대사능이 있는 미생물과 D-타가토오스 대사능이 결여된 미생물의 유전자군 비교 결과를 나타낸 도이다. 도 4은 D-타가토오스 대사능 관련 도 5는 도 6는 도 7은 도 8은 pET-22b(+)- 도 9는 도 10은 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자가 도입된 재조합 균주를 글루코오스 또는 D-갈락토오스 포함 배지에서 배양하고, 세포성장을 확인한 결과를 나타낸 도이다. 도 11은 세포성장 기반 초고속 탐색법을 이용하여 당이성화 효소 라이브러리 변이체의 균체 성장곡선을 나타낸 도이다. 도 12은 세포성장 기반 초고속 탐색법을 이용하여 선별된 당이성화효소 변이효소의 D-galactose 에 대한 활성을 야생형과 비교한 결과를 나타낸 도이다. 본 명세서에서 달리 정의되지 않은 용어들은 본 발명이 속하는 기술분야에서 통상적으로 사용되는 의미를 갖는 것이다. 본 발명은 1) 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계; 2) 제1당 대사능 및 제2당 이성화능이 결여된 균주에 상기 1) 단계에서 선별된 효소를 암호화하는 유전자를 도입하여 제1당 대사성 재조합 균주를 제조하는 단계; 3) 제2당 이성화 효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계; 4) 상기 3) 단계에서 수득된 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계에서 제조된 제1당 대사성 재조합 균주에 도입하여 균주 라이브러리를 얻는 단계; 및 5) 제2당을 유일 탄소원으로 포함하는 제한 배지에서 상기 4) 단계에서 수득된 라이브러리의 균주를 배양하고 균의 성장 및 생장속도를 확인하는 단계; 를 포함하며, 상기 제1당과 제2당은 상호 이성질체인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 및 제1당 대사능을 갖는 변이체의 초고속 스크리닝 방법을 제공한다. 본 발명의 스크리닝 방법을 통해 수득되는 변이체는 제2당 이성화능 및 제1당 대사능이 가능한 당 대사 경로가 새롭게 구축된 변이체이며, 새롭게 구축된 당 대사 경로에 의하여 제2당을 제1당으로 이성화시킬 수 있고, 이성화를 통해 생산된 제1당을 효과적으로 대사할 수 있다. 따라서 본 발명의 변이체는 제2당을 유일 탄소원으로 포함하는 배지에서 배양되는 경우, 당 대사 경로가 변경되지 않은 야생형 균주와 비교하여 높은 생장을 나타낼 수 있으며, 생장능 비교를 통해 무작위 유전자 변이를 유발한 변이 제2당 이성화 효소 중 제2당을 효과적으로 이성화시킬 수 있는 효소를 포함하는 균주를 빠르게 선별해 낼 수 있다. 본 발명에 사용되는 상호 이성질체 관계에 있는 제1당 및 제2당은 당 분야에 알려진 통상의 당 이성질체를 모두 제한없이 포함할 수 있으나, 바람직하게는 제1당은 타가토오스 및 제2당은 갈락토오스인 제1조합; 제1당은 프럭토오스 및 제2당은 글루코오스인 제2조합; 제1당은 타가토오스 및 제2당은 프럭토오스인 제3조합; 제1당은 만노오스 및 제2당은 글루코오스인 제4조합; 제1당은 글루코오스 및 제2당은 갈락토오스인 제5조합; 및 제1당은 프럭토오스 및 제2당은 갈락토오스인 제6조합; 으로 이루어진 군에서 선택된 조합인 것을 특징으로 할 수 있다. 당 분야의 통상의 기술자는 목적으로 하는 변이 당이성화 효소를 수득하기 위하여 필요한 당 대사 경로를 구축하기에 적절한 상호 이성질체 관계의 제1당 및 제2당을 선택하여 사용할 수 있다. 예컨대 본원 발명의 일 실시예에서는 D-갈락토오스 (제2당) 를 D-타가토오스 (제1당)로 전환시킬 수 있는 새로운 변이 이성화 효소를 수득하기 위한 방법을 개시한다. 먼저 제1당인 D-타가토오스 대사능을 부여할 수 있는 효소인 D-타가토오스-1,6-비스포스페이트 알돌라아제를 선별하여 이를 암호화하는 유전자 당 분야의 통상의 기술자는 제 1단계의 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계를 수행함에 있어서, 목적으로 하는 제1당의 대사능과 관련된 필수적인 효소를 도출하기 위해 필요한 균주의 비교 및 유전체 분석을 당 분야에 알려진 공지의 방법을 통해 제한없이 수행할 수 있다. 상기 1) 및 2) 단계와 같이 제1당 대사에 필수적인 효소, 예컨대 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자가 도입되는 경우, 종래 D-타가토오스만이 탄소원으로 첨가된 배지에서 균체 성장이 일어나지 않았던 D-타가토오스 대사능이 결여된 균주에 새로운 당 대사 경로가 구축됨으로써, D-타가토오스 첨가 배지에서 균체 성장이 일어날 수 있게 된다. 상기 3) 단계는 제2당 이성화 효소를 코딩하는 효소, 예컨대 L-아라비노오스 이성화효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계이며, 무작위 유전자 변이란 변이유발 PCT인 error-prone PCR 법을 통해 정상 서열을 주형으로 임의의 변이를 유발하는 것을 말한다. 상기 유전자 변이 라이브러리는 무작위 변위를 통해 수득된 변이 유전자의 집단, 집합을 말한다. 상기 4) 단계는 상기 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계의 제1당 대사성 재조합 균주, 예컨대 D-타가토오스 대사성 균주에 도입하여 균주 라이브러리를 얻는 단계를 말하며, D-타가토오스 대사성 균주가 변이 유전자를 추가적으로 발현하고, 이를 통해 새로운 당 대사 경로를 구축한 균주의 라이브러리를 얻는 것을 의미한다. 유전자 변이 라이브러리의 개별 변이 유전자가 1종씩 2) 단계의 D-타가토오스 대사성 균주에 도입되어 발현될 수 있으며, 이러한 공정을 통해 다양한 변이 유전자가 새롭게 도입된 균주 라이브러리를 얻을 수 있다. 상기 5) 단계는 상기 균주 라이브러리의 균주를 제2당을 유일 탄소원으로 포함하는 배지에서 배양하여 균의 성장을 확인하는 단계이며, 균의 성장을 확인하여 해당 균주가 제2당 전환능을 획득하였는지 여부를 빠르게 확인할 수 있다. 특히 균체의 성장이 우수하게 나타나는 균주를 제2당 전환능이 획득된 균주로 선별할 수 있으며, 해당 균주에 도입된 3) 단계의 유전자 변이 라이브러리의 변이 유전자를 확인함으로써, 제2당 전환능을 부여하는 제2당 이성화능을 갖는 변이체 및 이에 도입된 변이 효소를 도출해 낼 수 있다. 따라서 본 발명은 1) 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계; 2) 제1당 대사능 및 제2당 이성화능이 결여된 균주에 상기 1) 단계에서 선별된 효소를 암호화하는 유전자를 도입하여 제1당 대사성 재조합 균주를 제조하는 단계; 3) 제2당 이성화 효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계; 4) 상기 3) 단계에서 수득된 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계에서 제조된 제1당 대사성 재조합 균주에 도입하여 균주 라이브러리를 얻는 단계; 5) 제2당을 유일 탄소원으로 포함하는 제한 배지에서 상기 4) 단계에서 수득된 라이브러리의 균주를 배양하고 높은 성장능을 갖는 균주를 선별하는 단계; 및 6) 상기 5) 단계에서 선별된 균주에 도입된 변이 효소를 확인하는 단계; 를 포함하며,상기 제1당과 제2당은 상호 이성질체인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 을 갖는 변이 당이성화 효소의 초고속 스크리닝 방법을 제공한다. 상기 방법의 각 단계는 앞서 기술된 제2당 이성화능 및 제1당 대사능을 갖는 변이체의 고속 스크리닝 방법에 대한 설명에 따라 수행될 수 있다. 본 발명에서 일 실시예로 개시한, 당이성화능을 갖는 L-아라비노오스 이성화효소 변이효소 및 D-갈락토오스 당이성화 변이체의 고속 스크리닝 방법은 “세포성장 기반 초고속 탐색법”을 기반으로 하는 것을 특징으로 한다. 본 발명의 세포성장 기반 초고속 탐색법은 다양한 후보 변이 유전자들을 플랫폼 균주에 도입하고, 이에 대한 세포 성장을 확인함으로써 이를 기초로 활성을 나타내는 변이 유전자를 빠르게 선별, 탐색할 수 있는 것을 의미한다. 보다 구체적으로 본 발명에서는 다양한 변이 L-아라비노오스 이성화효소 코딩 유전자의 라이브러리를 형성하고, 이를 D-갈락토오스 이성화능이 결여된 균주에 도입하고 D-갈락토오스 포함 제한 배지에서 균주를 배양함으로써 균주의 성장을 확인하는 방법을 통해, 수많은 변이 L-아라비노오스 이성화효소 코딩 유전자 중 D-갈락토오스 이성화능을 효과적으로 부여할 수 있는 특정 변이 유전자를 도출하여 D-갈락토오스 당이성화 변이효소로 이용할 수 있도록 한다. 또한 본 발명은 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자를 포함하는, 재조합 벡터를 제공한다. 또한 본 발명은 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자가 도입된, D-갈락토오스 이성화능을 갖는 재조합 균주를 제공한다. 상기 D-타가토오스-1,6-비스포스페이트 알돌라아제를 암호화하는 유전자는 타가토스 대사능이 있는 미생물로부터 유래된 유전자일 수 있으며, 바람직하게는 또한 본 발명의 L-아라비노오스 이성화효소를 코딩하는 유전자는 갈락토스 대사능이 있는 미생물로부터 유래된 유전자일 수 있으며, 바람직하게는 본 발명의 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자를 포함하는 재조합 벡터를 이용하면, D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자가 도입된, D-갈락토오스 대사능을 갖는 재조합 균주를 수득할 수 있으며, 상기 재조합 균주는 외래 유전자 도입으로 새로운 변형된 당 대사 경로가 구축되므로, 종래 이용할 수 없었던 당, 바람직하게는 D-갈락토오스를 이용하여 유용물질인 D-타가토오스를 생산할 수 있는 대사능을 새롭게 가질 수 있다. 이와 같은 유전자 도입에 따라 구축된 새로운 당 대사 경로를 도 1에 모식도로 나타내었다. 본 발명에서 "당 이성화" 및 "당 전환"은 상호 교환적으로 사용될 수 있다. 본 발명의 "벡터”는 외래 유전자를 균주에 도입하기 위한 전달체를 말하는 것으로 바람직하게는 플라스미드일 수 있다. 상기 플라스미드는 당 분야에 통상적으로 널리 이용되는 것을 제한없이 사용할 수 있으나, 구체적으로 도 4에 나타낸 계열지도를 갖는 플라스미드를 이용하여 본 발명의 재조합 균주는 외래 유전자인 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자의 도입에 의하여 D-갈락토오스 대사능을 새롭게 가지게 된 균주이며, 특히 D-갈락토오스 대사능을 가지고 있지 않은 균주에 유전자가 도입된 균주이다. D-갈락토오스 대사능이 결여된 균주는 그 종류를 특별히 제한하지 않으나, 아그로박터리엄(Agrobacterium), 아조스피릴엄(Azospirillum) 바실러스(Bacillus), 버크홀데리아(Burkholderia), 클로스트리디엄(Clostridium), 엔테로박터(Enterobacter), 엔트로코커스(Enterococcus), 에스케리키아(Escherichia), 플라보박터리엄(Flavobacterium), 클렙시엘라(Klebsiella), 락토바실러스(Lactobacillus), 로도박터(Rhodobacter), 지오바실러스 (Geobacillus) 및 살모넬라(Salmonella)로 이루어진 그룹으로부터 선택된 1종의 속에 속하는 것일 수 있으며, 특히, 재조합에 가장 일반적으로 사용되는 엔테로박터(Enterobacter)에 속하는 대장균( 상기 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자가 도입된, D-갈락토오스 대사능을 갖는 재조합 균주는 D-갈락토오스를 D-타가토오스로 전환시키고, 타카토오스를 glycolysis에 이용할 수 있는 재조합 균주로, D-갈락토오스 및 D-타가토오스 대사능 또는 자화능을 모두 갖는 균주인 것을 특징으로 할 수 있다. 또한 본 발명은 D-갈락토오스 비대사성 균주에 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L-아라비노오스 이성화효소를 코딩하는 유전자를 도입하는 단계; 를 포함하는 D-갈락토오스 이성화능을 갖는 재조합 균주의 제조방법을 제공한다. 본 발명에서 상기 2 종의 유전자를 숙주 유전자에 도입하는 방법으로는 특별히 한정하지는 않으나, 벡터에 상기 유전자를 삽입하여 전기충격(electrophoresis) 및 열 충격 방법(heat shock transformation method) 등을 사용해 재조합 미생물 내에 도입함으로써, 재조합 균주에서 함께 발현될 수 있도록 하는 것이 바람직하고, 이용될 수 있는 벡터의 구체적인 예는 도 4 및 도 8에 나타내었다. 균주에 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자를 도입하는 경우, 상기 2종 유전자의 도입은 동시에 또는 순차적으로 이루어질 수 있으며, 순차적으로 이용하는 경우 그 순서에는 제한이 없다. 또한 본 발명은 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자가 도입된, D-갈락토오스 대사능을 갖는 재조합 균주를 D-갈락토오스 포함 배지에서 배양하는 단계; 를 포함하는 D-타가토오스 생산 방법을 제공한다. 상기 방법에 따르면, 기존에 D-갈락토오스 대사능이 결여된 균주라도 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자를 도입하는 공정을 통해 D-갈락토오스를 대사할 수 있게 함으로써, D-갈락토오스를 기질로 하는 배지에서 유용물질인 D-타가토오스를 생산할 수 있도록 할 수 있다. 상기 D-갈락토오스 포함 배지는 재조합 균주의 생장에 부합하는 성분을 포함하는 배지일 수 있으며, 탄소원으로는 D-갈락토오스 및/또는 포도당만을 포함하고 있는 제한 배지일 수 있다. 또한 본 발명은 서열번호 2 내지 4로 이루어진 군에서 선택된 1종의 아미노산 서열로 표시되는, 당이성화능을 갖는 L-아라비노오스 이성화효소 변이효소를 제공한다. 상기 변이 효소는 세포성장기반 초고속 탐색법으로 스크리닝된 당이성화효소 변이체의 변이 아미노산 서열이며, 아라비노오스 이성화효소를 무작위 변이시키고, 이를 균주에 도입한 후, D-갈락토오스 포함 배지에서 재조합 균주를 배양하고 성장을 확인함으로써 도출된 효소이다. 본 발명의 당이성화능을 갖는 L-아라비노오스 이성화효소 변이효소는 당 이성화 효소의 기능을 동등하게 나타내는 한, 서열번호 2, 3 및 4로 표시되는 효소들과 각각 90 내지 99%, 바람직하게는 95 내지 97%, 더욱 바람직하게는 97 내지 99%의 상동성을 갖는 서열을 모두 제한없이 포함할 수 있다. 이하, 본 발명을 실시예에 의해 상세히 설명한다. 단, 하기 실시예는 본 발명을 예시하는 것일 뿐 본 발명의 내용이 하기 실시예에 한정되는 것은 아니다. 실시예 1: D-갈락토오스 대사능이 결여된 대장균의 확보 기질인 D-갈락토오스에 대해 대사능이 없는 숙주세포를 선별하고 이를 실험적으로 확인하였다. D-갈락토오스에 대사능이 없는 대장균 BL21 (DE3)균주의 유전체 분석을 수행하였으며, 그 결과 해당 균주에서는 탄수화물 대사 경로 중 D-갈락토오스 대사에 관여하는 효소를 암호화하는 도 2에 나타낸 바와 같이, 대장균 DH5α는 포도당과 D-갈락토오스를 탄소원으로 사용할 수 있는 반면, 대장균 BL21 (DE3) 는 D-갈락토오스 첨가 배지에서 균체 성장이 일어나지 않는 것을 실험을 통해 확인하였다. 실시예 2: D-타가토오스 대사에 필수적인 외래 유전자 탐색 D-타가토오스 대사에 필수적인 효소군 및 유전자를 탐색하기 위하여, D-타가토오스 대사능이 있는 미생물 도 3에 나타낸 바와 같이, D-타가토오스 대사능이 있는 미생물인 실시예 3: GatY 도입을 통한 형질전환체의 대사능 획득 확인 3.1 GatY 단백질의 발현 벡터 및 대장균 형질전환균체의 제조 상기 실시예 2를 통해 D-타가토오스 대사에 필수적일 것으로 예상된 3.2 대장균내의GatY 단백질의 발현 확인 상기 3.1에서 수득된 재조합 과발현벡터 pET-28a(+) GatY 단백질 발현을 유도시킨 균체의 흡광도 (600 nm) 1 에 해당하는 배양액을 1.5 ml E-tube에 옮긴 후 원심분리 후 배양 균체만을 회수하였으며, 100 μl의 1X SDS sample buffer로 균체를 현탁한 후 끓는 물에서 10분간 처리 후 12% SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis) 분석을 수행하고 그 결과를 도 5에 나타내었다. 도 5에 나타낸 바와 같이, GatY 단백질의 발현(30 kDa)이 확인되었으며, 3.3 D-타가토오스를 이용한 형질전환균의균체성장 확인 재조합 유전자의 발현에 따라 대장균 BL21(DE3) 숙주 세포가 D-타가토오스 대사능을 획득하였는지 여부를 확인하기 위하여, 상기 3.1 내지 3.2를 통해 수득한 형질전환된 신규 대장균 숙주세포 pET-28a(+)- gatY/ 도 6에 나타낸 바와 같이, 외래유전자인 gatY가 도입된 형질전환균은 D-타가토오스 포함 제한배지에서도 글루코스 포함 배지와 유사한 정도로 우수한 성장 및 비증식속도를 나타냈으며, 도 7에 나타낸 바와 같이, D-tagatose 1, 6- bisphosphate aldolase를 암호화하는 gatY 외래유전자가 과발현된 대장균 BL21(DE3) 숙주세포는 외래 단백질 발현에 따라 균체 성장이 원활하게 일어남을 확인하였다. 그러나 이는 0.5% D-갈락토오스를 유일 탄소원으로 하는 제한배지에서는 균체 성장을 나타내지 않았다. 따라서 D-갈락토오스 대사능을 부여하기 위해서는 추가적인 당 대사 경로의 변형이 필요함을 확인하였다. 실시예 4: D-갈락토오스 이성화능을 갖는 균주의 제조 당이성화효소 라이브러리 탐색을 위한 플랫폼 균주 및 스크리닝 시스템 개발을 위해 상기 실시예 3에서 제조된 숙주세포, 즉 D-갈락토오스 대사능이 결여되고, D-타가토오스에 대해서는 대사능을 갖는 균주에 당이성화효소 L-아라비노오스 이성화효소(arabinose isomerase) 단백질 발현이 일어나도록 하였다. 이를 위해 L-아라비노오스 이성화효소 단백질의 발현벡터를 제조하였다. 발현벡터를 제조하기 위하여 먼저 각각의 제한효소 서열 ( 유전자의 발현을 위해 대장균 형질전환체를 아래와 같이 제조하였다. 상기 D-갈락토오스 비대사능이 확인되고, D-타가토오스 대사능이 탑재된 대장균 pET-28a(+)-gatY/BL21(DE3) 균주에 도 8에 나타낸 재조합 발현 벡터를 도입하였다. 형질전환균을 선택적으로 배양하기 위하여 상기 발현벡터에 포함된 항생제 내성 유전자에 적합한 항생제인 엠피실린 (ampicillin)이 최종농도 100 μg/ml이 포함된 LB 고체배지에 도말하고 37℃에서 배양하여 싱글 콜로니를 형성하였다. 상기 형질전환체를 전배양 후 플라스크 내에서 엠피실린 (100 μg/ml)을 첨가한 LB에 1%의 전배양균을 접종하고 본배양하여 흡광도 (600 nm)가 0.4-0.6일 때, 최종 농도 1 mM의 IPTG (Isopropyl -D-1-thiogalactopyranoside)를 사용하여 araA 유전자의 발현을 37℃에서 6시간 동안 유도하였다. 단백질 발현을 유도시킨 균체의 흡광도 (600 nm) 1에 해당하는 배양액을 1.5 ml E-tube에 옮긴 후 원심분리하고 배양 균체만을 회수하였으며, 100 μl 의 1X SDS sample buffer로 균체를 현탁한 후 끓는 물에서 10분간 처리 후 12% SDS-PAGE (sodium dodecyl sulphate-polyacrylamide gel electrophoresis) 분석을 통해 대장균 내에서의 D-tagatose 1, 6-bisphosphate aldolase 효소를 암호화하는 gatY 유전자의 발현 (31 kDa) 및 L-arabinose isomerase 효소를 암호화하는 araA 유전자의 발현 (56 kDa)여부를 도 9와 같이 확인하였다. 도 9에 나타낸 바와 같이, 형질전환된 대장균에서는 D-tagatose 1, 6-bisphosphate aldolase 효소를 암호화하는 gatY 유전자 (31 kDa) 및 L-arabinose isomerase 효소를 암호화하는 araA 유전자(56 kDa)가 발현하였다. 제작된 세포성장기반 스크리닝 탐색법의 이성화 효소의 활성 대비 및 세포성장을 확인하기 위해, 상기 두 플라스미드 pET-22b(+)- 도 10에 나타낸 바와 같이, D-갈락토오스를 포함하는 배지에서 성장하지 못하는 대장균 BL21(DE3) 균주에 D-tagatose 1, 6-bisphosphate aldolase 효소를 암호화하는 gatY 유전자 및 L-arabinose isomerase 효소를 암호화하는 araA 유전자가 도입된 균주는 균체 성장이 일어났으며, 이성화 효소의 활성으로 D-갈락토오스를 유일 탄소원으로 이용할 수 있음을 확인하였다. 실시예 5: araA 유전자의 변이-유발을 통한, 당이성화 효소 변이체 선별 5.1 araA 유전자의 변이-유발 (error-prone) PCR 및 변이 다양성 확인 L-아라비노오스 이성화효소 araA 유전자의 변이를 유발하기 위해, PCR 무작이 돌연변이 키트 (Clontech, 미국)로 변이 PCR을 수행하였다. 변이-유발 PCR 라이브러리 DNA 50 ng을 플라스미드 pET-28a(+)-gatY가 포함된 대장균 BL21(DE3)에 형질전환하였으며, 0.05g/L의 프락토스와 0.45%의 D-갈락토오스가 포함된 제한(M9)배지에서 배양을 수행하였다. 그 후, 형성된 콜로니를 모아 플라스미드 정제 키트를 이용하여 플라스미드를 추출하였다. 그 중 일부 플라스미드 8개의 염기서열을 분석하였으며, 라이브러리의 유전자 다양성을 확인하였다. 5.2 세포성장기반 초고속 탐색법을 이용한 당이성화효소변이체 선발 상기 다양성이 확인된 당이성화효소 라이브러리 유전자 변이 아라비노오스 이성화효소 유전자 pET-22b(+)-araA library DNA가 형질전환된 pET-22b(+)-araA library DNA 및 변이를 유발하지 않은 pET-28a(+)-gatY/BL21(DE3)를 탄소원으로 0.05 g/L 프락토오스와 0.45 g/L D-갈락토오스, 및 최종 농도 0.2 mM IPTG가 포함된 제한 (M9)배지에서 배양하고 균체성장을 확인하였으며, 그 결과를 도 11에 나타내었다. 도 11에 나타낸 바와 같이, 변이 라이브러리 유전자의 단백질 발현이 다양하게 나타남에 따라 균체 성장의 수준 및 비증식속도는 다르게 나타남을 확인하였으며, 이를 통해 araA 유전자의 도입 및 이의 변이에 따라 D-갈락토오스를 탄소원으로 이용할 수 있는 능력이 각각 상이하게 나타날 수 있음을 확인하였다. 5.3. 당이성화효소 변이효소의 D-갈락토오스 이용능 검증 상기 5.2의 세포성장기반 스크리닝 탐색법으로 선발된 당이성화효소 변이체 및 야생형으로부터 하기와 같이 플라스미드를 분리하였다: pET-22b(+)-ECAI H17R/R159S/V168A, pET-22b(+)-ECAI E22D/M95L/H157L, 및 pET-22b(+)-ECAI-V368A/E493D. 또한 분리된 플라스미드 유래의 서열분석을 통하여 스크리닝 탐색법으로 선발된 변이체들의 변이 아미노산 서열을 분석하였으며, 이를 각각 서열번호 2 내지 4에 나타내었다. 야생형 ECAI 의 아미노산 서열은 서열번호 1에 나타내었다. 추가적으로 Ni2+ 컬럼 크로마토그래피를 수행하여 변이효소를 순수분리하여 ECAI 야생형 효소와의 활성비교를 수행하였다. 즉, 숙주 균주 (Host), ECAI 야생형 및 당이성화 효소 변이체들의 D-갈락토오스에 대한 활성을 측정, 비교하였으며, 이를 도 12에 나타내었다. 도 12에 나타낸 바와 같이, pET-22b(+)-ECAI H17R/R159S/V168A 유래 변이효소 (서열번호 2), pET-22b(+)-ECAI E22D/M95L/H157L 유래 변이효소 (서열번호 3), 및 pET-22b(+)-ECAI-V368A/E493D 유래 변이효소 (서열번호 4) 의 경우 D-갈락토오스에 대한 활성이 최대 80% 배 증가하였음을 확인하였다. 이와 같은 결과를 통해, L- 아라비노오스 이성화효소의 무작위적인 변이를 유도하고 라이브러리를 형성한 후, 이를 균주에 도입하여 균주 라이브러리를 형성하고 균주 성장을 통한 D-갈락토오스 대사능을 확인하는 경우, D-갈락토오스 대사능을 부여할 수 있는 특정 변이 효소 및 당이성화 효소 변이체를 빠르게 선별할 수 있음을 확인하였다. 비록 본 발명이 상기에 언급된 바람직한 실시예로서 설명되었으나, 발명의 요지와 범위로부터 벗어남이 없이 다양한 수정이나 변형을 하는 것이 가능하다. 또한 첨부된 청구 범위는 본 발명의 요지에 속하는 이러한 수정이나 변형을 포함한다. The present invention relates to a recombinant strain having a modified sugar metabolic pathway resulting from the introduction of an enzyme derived from a different strain to a strain, and a high throughput screening method for various variant strains and mutant enzymes by which useful materials can be obtained or which can produce useful materials. The use of a recombinant vector and a strain according to the present invention not only can construct a new metabolic pathway in a strain to effectively obtain D-tagatose from D-galactose, but also can introduce randomly modified sugar isomerases and then allow D-galactose isomerase mutants to be rapidly screened by conducting a cell growth-based high-throughput screening method. 1) 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계; 2) 제1당 대사능 및 제2당 이성화능이 결여된 균주에 상기 1) 단계에서 선별된 효소를 암호화하는 유전자를 도입하여 제1당 대사성 재조합 균주를 제조하는 단계; 3) 제2당 이성화 효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계; 4) 상기 3) 단계에서 수득된 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계에서 제조된 제1당 대사성 재조합 균주에 도입하여 균주 라이브러리를 얻는 단계; 및 5) 제2당을 유일 탄소원으로 포함하는 제한 배지에서 상기 4) 단계에서 수득된 라이브러리의 균주를 배양하고 균의 성장 및 생장속도를 확인하는 단계; 를 포함하며, 상기 제1당과 제2당은 상호 이성질체인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 및 제1당 대사능을 갖는 변이체의 초고속 스크리닝 방법. 제1항에 있어서, 제1당 및 제2당은 제1당은 타가토오스 및 제2당은 갈락토오스인 제1조합; 제1당은 프럭토오스 및 제2당은 글루코오스인 제2조합; 제1당은 타가토오스 및 제2당은 프럭토오스인 제3조합; 제1당은 만노오스 및 제2당은 글루코오스인 제4조합; 제1당은 글루코오스 및 제2당은 갈락토오스인 제5조합; 및 제1당은 프럭토오스 및 제2당은 갈락토오스인 제6조합; 으로 이루어진 군에서 선택된 조합인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 및 제1당 대사능을 갖는 변이체의 초고속 스크리닝 방법. 제2항에 있어서, 제1당은 타가토오스 및 제2당은 갈락토오스이며, 제1단계에서 선별된 효소는 D-타가토오스-1,6-비스포스페이트 알돌라아제, 제3단계의 이성화 효소는 L- 아라비노오스 이성화효소인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 및 제1당 대사능을 갖는 변이체의 초고속 스크리닝 방법. 제1항에 있어서, 상기 3) 단계의 무작위 유전자 변이는 변이-유발 (error-prone) PCR을 통해 수행되는 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 및 제1당 대사능을 갖는 변이체의 초고속 스크리닝 방법. 1) 제1당 대사 균주와 제1당 대사능이 결여된 균주의 유전체 분석을 통해 제1당 대사에 필수적인 효소를 선별하는 단계; 2) 제1당 대사능 및 제2당 이성화능이 결여된 균주에 상기 1) 단계에서 선별된 효소를 암호화하는 유전자를 도입하여 제1당 대사성 재조합 균주를 제조하는 단계; 3) 제2당 이성화 효소를 코딩하는 유전자에 무작위 유전자 변이를 유발하여 유전자 변이 라이브러리를 수득하는 단계; 4) 상기 3) 단계에서 수득된 유전자 변이 라이브러리의 변이 유전자를 상기 2) 단계에서 제조된 제1당 대사성 재조합 균주에 도입하여 균주 라이브러리를 얻는 단계; 5) 제2당을 유일 탄소원으로 포함하는 제한 배지에서 상기 4) 단계에서 수득된 라이브러리의 균주를 배양하고 높은 성장능을 갖는 균주를 선별하는 단계; 및 6) 상기 5) 단계에서 선별된 균주에 도입된 변이 효소를 확인하는 단계; 를 포함하며, 상기 제1당과 제2당은 상호 이성질체인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 을 갖는 변이 당이성화 효소의 초고속 스크리닝 방법. 제5항에 있어서, 제1당 및 제2당은 제1당은 타가토오스 및 제2당은 갈락토오스인 제1조합; 제1당은 프럭토오스 및 제2당은 글루코오스인 제2조합; 제1당은 타가토오스 및 제2당은 프럭토오스인 제3조합; 제1당은 만노오스 및 제2당은 글루코오스인 제4조합; 제1당은 글루코오스 및 제2당은 갈락토오스인 제5조합; 및 제1당은 프럭토오스 및 제2당은 갈락토오스인 제6조합; 으로 이루어진 군에서 선택된 조합인 것을 특징으로 하는, 제2당 이성화능 (isomerization activity) 을 갖는 변이 당이성화 효소의 초고속 스크리닝 방법. D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자를 포함하는, 재조합 벡터. 제7항에 있어서, 상기 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자는 제7항에 있어서, 상기 L- 아라비노오스 이성화효소를 코딩하는 유전자는 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자가 도입된, D-갈락토오스 이성화능을 갖는 재조합 균주. D-갈락토오스 비대사성 균주에 D-타가토오스-1,6-비스포스페이트 알돌라아제를 코딩하는 유전자 및 L- 아라비노오스 이성화효소를 코딩하는 유전자를 도입하는 단계; 를 포함하는 D-갈락토오스 이성화능을 갖는 재조합 균주의 제조방법. 제10항의 재조합 균주를 D-갈락토오스 포함배지에서 배양하는 단계; 를 포함하는 D-타가토오스 생산 방법. 서열번호 2 내지 4로 이루어진 군에서 선택된 1종의 아미노산 서열로 표시되는, D-갈락토오스 이성화능을 갖는 L-아라비노오스 이성화효소 변이효소.