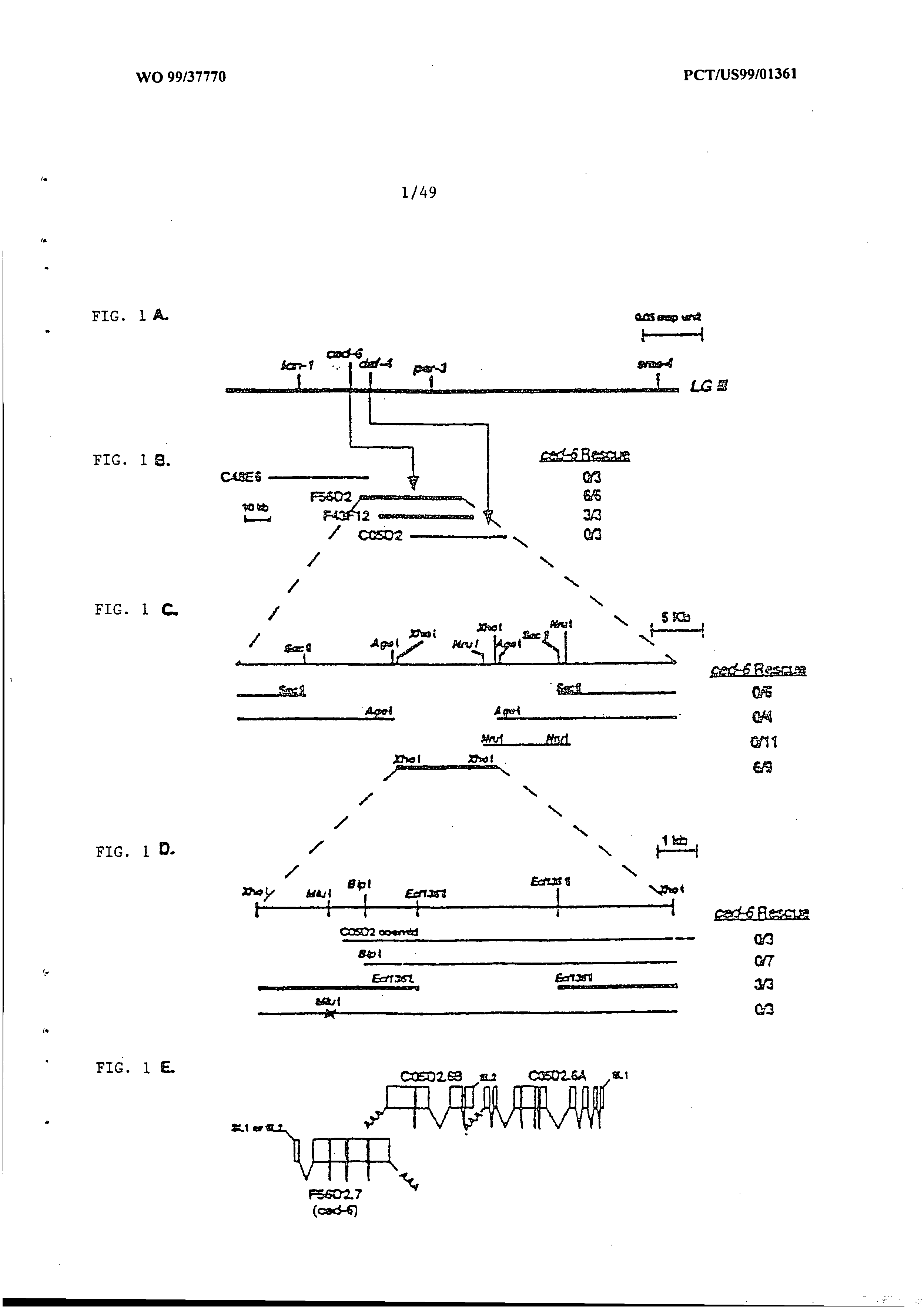
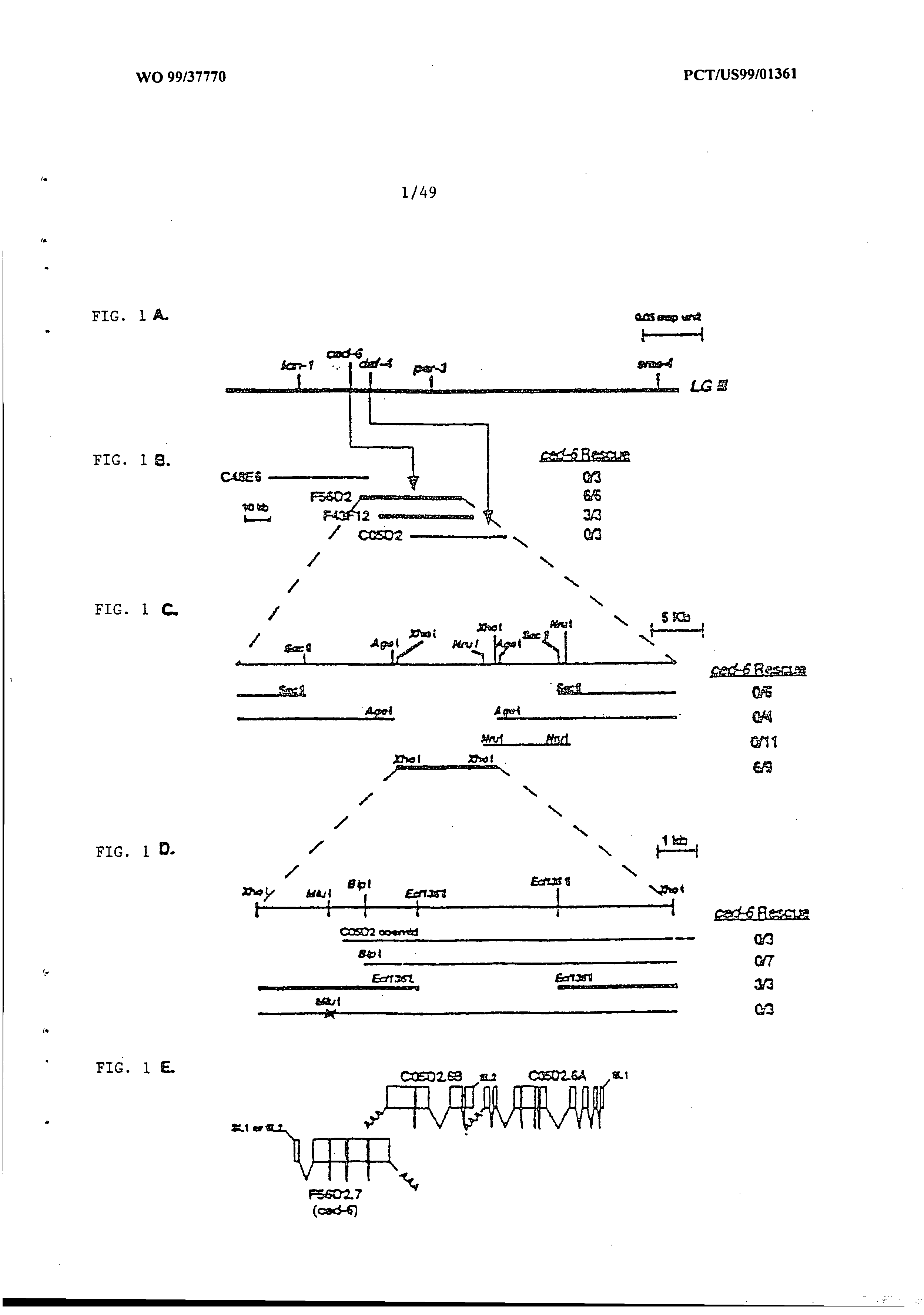
Phagocytosis genes and uses thereof
PHAGOCYTOSIS GENES AND USES THEREOF BACKGROUND TO THE INVENTION Phagocytosis or engulfment, is a specialized form of endocytosis through which eukaryotes take up very large particles, or even whole cells. It is a fundamental biological process conserved from single-cell organisms, such as amoebae to mammals (Metchnikoff, E. 1891), Lectures on the comparative pathology of inflammation; delivered at the Pasteur Institute, 1891, 1968 Edition (New York: Dover Publication)). Initially used for the dual purpose of feeding and defence, phagocytosis evolved, following the emergence of mesoderm, into a mechanism used to protect the host against invading organisms and to clear up foreign particles and cell debris (Metchnikoff, 1891). Recently, the significance of phagocytosis has been extended due to its role in eliminating cells undergoing programmed cell death (apoptosis). Since apoptosis has been implicated in a number of human diseases elucidation of the regulation of this phagocytosis is highly desirable since it may lead to a new route of therapeutic intervention in these diseases. Accordingly, a need exists to isolate a gene and protein that regulate phagocytosis. A further need exists for therapeutic treatment for diseases related to phagocytosis of apoptotic cells. SUMMARY OF THE INVENTION Genetic studies in C. elegans have identified over a dozen genes that function in programmed cell death. The present inventors have used the positional method to clone and have functionally characterized the C. elegans gene CED-6. It is shown that the CED-6 protein contains a phosphotyresine binding domain and several potential SH3 binding sites. It is further demonstrated that CED-6 acts within engulfing cells, and functions to promote the removal of both early and persistent cell corpses. Overexpression of CED-6 can partially suppress the engulfment defect of both CED1 and CED-7, suggesting that CED-6 functions downstream of these two genes. CED-6 acts as an adaptor molecule in a signal transduction pathway that mediates the engulfment of apoptotic cells in C. elegans. The present inventors have also identified isolated and characterized human CED6 homologue including a splice variant thereof, which it is shown is involved in a similar process in mammalian cells. In one embodiment the invention provides, in isolated form, a protein which is the CED-6 protein of C. elegans or a protein which has equivalent function thereto and human homologues of the protein, hereinafter referred to as hlCED-6, h2CED-6, and h3CED-6. In another embodiment the invention provides a functional fragment of CED-6, hlCED-6, h2CED-6 and h3CED-6, for example, a fragment corresponding to the phosphotyrosine binding domain and/or the proline/serine rich region. In yet a further embodiment the invention provides an isolated nucleic acid encoding CED-6 and human homologues of CED-6, as well as nucleic acid encoding functional fragments of CED-6, hl CED-6, h2-CED-6 and h3-CED-6 as described above. The invention further provides in one embodiment nucleic acid which is antisense to any of the nucleic acids described above or which is capable of hybridizing to any of the nucleic acids described above under conditions of low, medium or high stringency or portions of fragments thereof. The invention further provides in another embodiment expression vectors comprising nucleic acid encoding CED-6, hl CED-6, h2CED-6, h3CED-6 or encoding functional fragments of said proteins as above. The invention further provides in yet another embodiment mammalian cell-lines transfected with one or more nucleic acids encoding CED-6, hl CED-6, h2CED-6, and/or h3CED-6. The invention further provides in one embodiment assay methods using the proteins, nucleic acids and transfected cells described above to identify compounds which enhance or inhibit the signal transduction pathway in which CED-6, hl CED-6, h2CED-6, and/or h3CED-6 participate. The invention further provides in another embodiment assay methods using the transfected cells described above to identify compounds which enhance or inhibit the expression of the CED-6, hlCED-6, h2CED-6 or h3CED-6 genes. The invention further provides antibodies which react with an epitope of CED-6, hl CED-6, h2CED-6, and/or h3CED-6. The invention further provides in yet another embodiment a method of treating diseases the etiology of which may be attributed to failure of engulfment of apoptotic or other diseased cells such as inflammation autoimmune disease or cancer by administering to a patient one or more of the aforesaid proteins or nucleic acids or compounds which are enhancers of CED-6, hl CED-6, h2CED-6 or h3CED-6. The invention further provides in one embodiment a method of treating diseases which would benefit from a reduction in the engulfment of apoptotic cells, such as, neurodegenerative diseases, stroke, or sickle-cell anaemia, by administering one or more of the aforesaid proteins, nucleic acids or compounds which are inhibitors of CED-6, hlCED-6, h2CED-6, or h3CED-6. The invention further provides in one embodiment a method of diagnosis of a human or animal disease using a nucleic acid encoding CED-6, hl CED-6, h2CED-6 or h3CED-6 or the complement thereof or an antibody to CED-6, hlCED-6, h2CED-6 or h3CED-6 to detect a genetic defect. The invention further provides in yet another embodiment a method of identifying proteins which interact with CED-6, hl CED-6, h2CED-6 or h3CED-6 in the signal transduction pathway in which those proteins participate. The invention further provides in another embodiment a fusion protein in which CED-6, hl CED-6, h2CED-6 or h3CED-6 or a functional fragment thereof such as the phosphotyrosine binding domain or serine proline rich region, is fused to another protein such as an epitope tag or product of a reporter gene. The invention further provides in yet another embodiment a method of determining whether a compound is an enhancer or inhibitor of the signal transduction pathway in which CED-6 participates by observing the effect of the compound on C. elegans worms having altered CED-6 expression. BRIEF DESCiLIPTION OF THE DRAWINGS Figure 1A - 1E are schematic representation of the CED-6 Locus. Figure 1A Genetic map of CED-6. CED-6 and some genes close to and also used to map CED-6 are shown. Figure 1B Cosmid rescue. Transgenic animals carrying cosmids or subcloned DNA fragments (see C, D) were examined for cell corpses on three fold embryos. Those who gave embryos with partial or no cell corpses were counted as rescuing transgenic lines. Four out of tested thirteen cosmids are shown. Rescuing fragments are bold. Number represents # rescuing lines/# lines tested. Figure 1C Subcloning of F56D2 cosmid and rescue. Restriction map of the CED-6 region is shown on the top. In the middle, several restriction fragments were tested for their ability to rescue the engulfment defect caused by CED-6(n1813). Figure 1D Subcloning of I 0 kb Xho I fragment and rescue. Restriction map ofXho I fragment is shown on the top. In the middle mutations made on the Xho I fragment and their rescuing ability are shown. An X indicates a frameshift mutation (see Experimental Procedures for details). Figure 1E Transcripts on Xho I fragments. Intron/exon structure of the transcripts on Xho I fragment region. Boxes: cons; V symbol: introns. AAA: poly(A) tail. RT-PCR products ofh' end of F56D2.7 contain both SLI and SL2. FigLtrcs 2A and B shows that F56D2.7 Encodes CED-6. Figure 2A shows the full-length eDNA (SEQ ID NO: 1.) and amino acid (SEQ ID NO: 2) of C. elegaas CED-6. Doub]* underline shows thc nucleic, acid (SEQ 1D NO: 3) and amino acid scxlttenc¢ (SEQ ID NO: 4) ofphosphotyrosinc binding (PTB) domain, singlc underline indicates the nucleic acid (SEQ ID NO: 5) and the ,amino acid (SEQ ID NO: 6) sequence of the prolineJscfinv rich region, Dashed underline indicates charaeA region. Star identifies the prolines in thc PxxP signature seqncncc, empty triangles the charged rcsidues within the dashed region. Shaded box indicates polyadcnylation signal. Both SLI and SL2 conld be added to transplicing accepter site. The single base pair deletion identified in CED-6(n1813) is shown. Figure 2B Southern blot which rcvcalcd a RFLP on 4.1 kb fragment from CED-6 (n209.5). Xho I probe identified an allclc-specihc RFLP in UED-6(n2095) that affect a 4.1 kb Hind Ill fragment containing, F56D2.7. On ti 1 e right bottom the genomic fragments digested by Hind Ill on the Xho l fragment region is shown. On the right top X/to 1 fragment and three genes covered on this xcgion, Thr Hbtd Ill fragments, 4.lkb, 0.4 kb and 9.9 kb lhat should be lighted up on thc Southern blot are indicated. On the left genomic DNA isolated independently from wild-type N2, CED-6(n 1813) and CED-6(n2095) wcrc p[obcd with riP-labeled Xho I fragmmlt, n2095 allele showed the missing of the 4.1 kb ['ragmcnt and the extra 2.1 kb fragment, 0.4 kb fragments wcrc not affected in both alleles (data on a separate gel, not shown here), Figure 3A-C show that CED-6 Contains a Phosphatyrosinv Binding Domain. Figure 3A shows that alignment ofCED-6 PTB (SEQ ID NO: 4) with other PTB domain. The PTB domain alignment was based on the NMR structure of She protein. Black boxes indicate identical amino acid showed by >50% of sequences. Grey boxes indicate similar amino acid showed by >50% o/" sequences. For this purpose, the following scts ofmnino acids are considcrcd similar: O, A, C, S, T; B, D, Q, N; R, K, I-l; V, M, L, I; F, Y, W indicate the a AMENDED SHEET helices suggested by the NMR structure of Shc, and [3 the 13 sheats. Invariant residues (found in all sequences shown) are highlighted by star, "*". Figure 3B shows the comparison of CED-6 to other PTB domain containing proteins. Proline rich regions and charged regions next to PTB domains and other regions. PTB domains were compared in the percentage of identity. Figure 3C shows the evolution tree of the PTB domains. The alignment from (A) was displayed using Seqlab package in GCG program, and the evolution tree was grown graphically. Figure 4 shows results of the Genetic Mosaic Analysis for CED-6 (table at bottom) and Cell lineage of C. elegans (top). The descendence of both germline and somatic sheath cells are illustrated. Body wall muscles cells which were used to determine the loss of the duplication were also illustrated. The solid square indicates the duplication loss in germ cells, and the solid square indicates the duplication loss in the somatic sheath cells. The black arrow indicates the somatic sheath cell with the enlarged nucleoli in the distal arm of the anterior gonad. The white arrow indicates the cell corpses accumulated in the proximal arm of anterior gonad. Figure 5A-D provide results that showed that heat-shock overexpression of CED-6 cDNA rescued the engulfment defect in both soma and germline. Figure 5A shows the cell death during the embryonic development. Shaded box is a histogragh indicating the number of dying cells every 50 minutes during the embryonic development. The arrows indicates the timing of heat shock and the timing to observe the engulfment phenotype. Figure 5B shows the overexpression of CED-6 cDNA promotes the engulfment at both the early and the late stage of cell death. Transgenic animals carrying the transgene, CED-6 cDNA driven by heat shock promoter were treated with heat before the cell death occurred at the indicated time. Cell Corpses in the head of young L 1 larvae were examined. The animals without the heat treatment were also examined. Other control experiments included N2, CED-6(n1813) with or without heat treatment, and CED-6(n1813) carrying lacZ transgene treated with heat. The solid circles indicate the experiments with the heat shock after the formation of cell Corpses, and the empty circles with the heat shock before the cell death took place and the experiments without heat shock. Figure 5C shows the overexpression of CED-6 cDNA rescue the engulfment defect in germline. The arrow indicates the timing for a heat shock when transgenic animals were at the development stage of the 24 hours after the L4 molt. Cell corpses were examined at the several time points between the time of heat shock and the 60 hours after the heat shock. Figure 5D shows the overexpression of CED-6 cDNA promotes the engulfment many hours after the formation of the cell corpses in germline. Adult transgenic animals were treated with heat as indicated. Cell corpses were examined in one gonad arm 12 hours after the heat shock. Control experiments including N2, and CED-6(n1813) are indicated in (C). Figure 6 presents results that show overexpression of CED-6 partially suppresses the engulfment defect of both CED-1 and CED-7 during embryonic development CED-6 was overexpressed at the genetic background of three alleles of both CED-1 and CED-7. The timing for the heat shock and the timing for the examination of cell corpses are illustrated in figure 5A. Animals with each genetic background were treated with heat before the cell death occurred or without the heat treatment. Cell corpses were examined in head of young L1 larvae. LacZ was also expressed in the each genetic background. Each mutant was also treated with heat shock to examine the effect of heat on the expression of cell corpses. Figure 7 is a model of the epistatic pathway for the engulfment genes overexpression of CED-6 did not have an obvious effect on the cell corpses expression on CED-2, 5 and 10 but on CED-1 and CED-7. We propose that CED-6 might act downstream of both CED-1 and CED-7. And CED-2, 5 and either act in the different pathway or act downstream of CED-6. Figure 8 is a flow chart illustrating a Xho I fragment from F56 cosmid rescues the CED-6 engulfment defect. Figure 9A-B are schematics that illustrate that the C05D2.7 construct is CED-6. Figure 9A shows the restriction Map of Xho I fragment and rescue. Figure 9B shows the transcripts. Figure 10 is a bar graph illustrating that the over-expression of CED-6 rescues the engulfment defect of the CED-6 mutant. Figure 11 contains graphs illustrating that the over-expression of CED-6 rescues the engulfment defect of CED-6 mutant during embryonic development. Figure 12 is a bar graph illustrating that CED-6 may also promote the engulfment of persisting corpses. Figure 13 shows that CED-6 promotes the engulfment of persistent cell corpses and probably acts within engulfing cells. Figure 14 is a schematic that shows that CED-6 may be an adaptor protein acting in signal transduction pathway. Figure 15 shows graphs which indicate that over-expression of CED-6 rescues the engulfment defect in the adult gonad, and CED-6 might act in somatic sheath cells. Figure 16 illustrates that over-expression of CED-6 partially suppresses the engulfment defect of CED1 mutants. Figure 17 shows that the over-expression of CED-6 cDNA suppresses the engulfment defect of CED-7 mutants. Figure 18 shows consensus DNA sequence (SEQ ID NO: 7) ofhlCED-6 (2416bp) with start and stop codon in bold and alternatively spliced sequence underlined. Figure 19 shows DNA sequence (SEQ ID NO: 13) ofh2CED-6 (altemative splice) with start and stop codons in bold. Figure 20 shows the amino acid sequence (SEQ ID NO: 8) ofhlCED-6 with alternatively spliced region underlined. Figure 21 shows the amino acid sequence (SEQ ID NO: 14) of h2CED-6 (alternative splice). Figure 22 shows hlCED-6 cDNA (SEQ ID NO: 7) and hlCED-6 (SEQ ID NO: 8) amino acid sequence with PTB domain nucleic (SEQ ID NO: 9) and amino acid (SEQ ID NO: 10) sequences, charged region, and proline/serine rich nucleic acid (SEQ ID NO: 11) and amino acid (SEQ ID NO: 12) sequences indicated. Figure 23 shows an alignment of CED-6 and h 1CED-6. Figure 24 shows an alignment of regions of 47.5% and 31.6% identity, respectively. Figure 25A Human Multiple Tissue Northern Blot (MTN), Figure 25B shows a Human Multiple Tissue Northern (MTN) Blot II, and Figure 25C shows a Human Cancer Cell Line Multiple Tissue Northern (MTN TM) Blot. The expression pattern ofhlCED-6 in normal human tissues and cancer cell lines by Northern blotting is shown. Figure 26 is a map of plasmid pGA3015 in which a CED-6 fragment is cloned as a C-terminal fusion to GFP. Figure 27 is a map of plasmid pGA3064 with CED-6 cloned as a Cterminal fusion of GFP. Figure 28A-28F is a DNA alignment (Genework) of sequenced hbc3123 EST clone, the PCR fragment I isolated from a cDNA library, and three EST sequences identified using the PCR fragment, hbc3123 EST clone was sequenced and analyzed. The three EST clones were identified through searching the Genbank using the isolated PCR fragment. Figure 29 shows the amino acid sequence (SEQ ID NO: 16) of the human h3 CED-6, as compared to hlCED-6 (SEQ ID NO: 8). Figures 30A-B show the nucleic acid sequence (SEQ ID NO: 15) that encodes human h3 CED-6, as compared to hlCED-6 (SEQ ID NO: 7). Figures 31A-B show that overexpression of h3CED-6 rescue an engulfment defect. Figures 31A shows overexpression of hCED-6 rescued the engulfment defect of CED-6(ni 813) embryos. Embryos laid by transgenic mothers were heat- shocked before the wave of embryonic cell death, and scored for the numbers of persistent cell corpses in head of L 1 larvae. Each dot represents one animal. Figure 31B shows overexpression of hCED-6 rescued the germ cell engulfment defect of CED-6(nI813) animals. Transgenic animals were heat-shocked 36 hours after L4/adult molt, and germ cell corpses were scored 12 hours after heat shock. The number of animals scored is indicated on the top of each bar. Figure 32A-J shows the nucleic acid sequence comparison among ESTs, CED-6, hCED-6, and a consensus construction of 2416 bp consensus sequence was done by using sequence information obtained from EST RACE & colony hybridization. Seq was compiled by using aa1599394 as template and primers as indicated in multiple alignment. Rcc stands for the reverse complement. Both CED-6 and hCED-6 are indicated above the multiple alignment pGA101 was picked up by colony hybridization. DETAILED DESCRIPTION OF THE INVENTION cDNAs encoding the alternative splice h2CED-6 and the additional sequence required to constitute h3CED-6 from h2CED-6 have been deposited at the Belgian Coordinated Collections of Microorganisms (BCCM) at Laboratorium voor Moleculaire Biologic - plasmidencollective (LMBP), Universiteit Gent, K.L. Ledeganckstraat 35, B 9000, Gent, Belgium in accordance with the Budapest Treaty on 8th June 1998 and have been accorded the Accession Nos LMBP 3868 and LMBP 3869, respectively. Primers which will assist in obtaining the relevant inserts from these deposits are shown in Example 14. AMINO ACID AND NUCLEOTIDE SEQUENCES Nucl c acid sequcn'cc ofC. ¢legans CED-6. (eg, Figure 2A) ..... Amino Acid sequence of (Z elegant CED-6 (e.g., Figme 2A) Nucleotide sequence t,'ncoding PTB domain of C. elegant" CED-6 (e.g., Figure 2A) , Hi Amino acid sequence oflq'B domain or" €. elegmas CED-6 (e.g., Figure 2A) Nuclcotidc seqm--nce encoding proliae./scrin¢ rich region of C. etegans CED-6 (e.g.. Figure 2A) al ,m Amino acid sequence of prolinJsta-in¢ rich region of C elegans CED-6 (e.g., Figure 2A) Nuoleotidt: sequence that encodes hlCEIY6 (e.g., Figure 22,Fibrurc 18) ,i , , - Amino acid sequence ofhlCED-6 (€.g., Figure 20 and Figure 22) m, Nuolcotid¢ sequence encoding, FIB domain of hlCED-6 (o.g.,Figurc 22) Amino acid sequ¢-ac¢ encoding PTB domain or h1CED-6 (e.g., Figure 2z.) ,m= Nucleic acid sequence aL encodes the proline/serinc rich region of hlCED-6 (e.g., Figure 22) rnino ac.id sequence of the prolinc/sc inc rich region of E1CEI)-6 (e.g., Figure 22) m ,, ,, i Nuolcotide sequence that encodes h2C'ED.6 (e.g., Figure 19) i = , , , ii ii _ A m ino acid sequence of h2CED-6 (e.g., Fit,,xn'c 2 l) iJ Nucleotide seq,aence encoding h3C -6 (e.g. Figure 30A-B) Amino acid sequence of h3CED-6 (e.g. Figure 29) AMENDED SHEET C. ELEGANS CED-6 Programmed cell death has traditionally been divided into two distinct, sequential processes: cell killing, and the removal of dead cells. However, these two events are very closely linked. In vivo, cells that present an apoptotic morphology are usually already engulfed by other cells (Wyllie A. H. et al., 1980 Int. Rev. Cytol 68, 251-306; Lockshin R.A. (1981) Cell Death in Biology and Pathology, R.A. Lockshin and I.D. Browen, eds. (London: Clapman and Hall), pp79-122; Duvall and Wyllie (1986). Immunol Today 7 pp 115-119; Robertson and Thompson (1982) J. Embryol. Exp. Morph. 67 pp 89-100; Hedgecock et al (1983) Science 222, 1277-1279; Ellis et al (1991) Genetics 129 pp 79-94;). Engulfment is also a swift and efficient process in the nematode Caenorhabditis elegans : dying cells are engulfed and completely removed by their neighboring cells within an hour (Sulston and Horvitz, (1977); Dev. Biol 56 pp 110-156; Robertson and Thomson, 1982). The engulfment is not necessarily by professional phagocytes. Rapid engulfment of apoptotic cells is important, as it prevents dying cells from releasing potentially harmful contents during their lysis, which could damage surrounding tissue and result in an inflammatory response (Duvall et al., (1985) Immunology 56 pp 351-358; Savill et al., (1989) J. Clin. Invest. 83 pp 865-875; Grigg et al., (1991) Lancet 358 pp 720-722; Savill et al., (1993) Irnmunol. Today 14, pp 131-136). The nematode C. elegans has been used extensively for the study of programmed cell death (reviewed by Hengartner, (1997) Cell Death in C. elegans II, Plain View, Cold Spring Harbour Laboratory Press, pp 383-415). Genetic studies have identified over a dozen genes that function in the regulation and execution of apoptosis in C. elegans. Six genes - CED-1, CED-2, CED-5, CED-6, CED-7, and CED-10 - function in the engulfment of all dying cells (Hedgecock et al., 1983; Ellis et al., 1991; Horvitz et al., (1994) Cold Spring Harbour Symp. Quant Biol (1994) 59: pp 377-385). In animals mutant for any one of these genes, many apoptotic cells fail to be engulfed and persist for many hours as highly refractile disks that can be readily identified under differential interference contrast (DIC) optics (Hedgecock et al., 1983; Ellis et al., 1991). None of the six engulfment genes is absolutely essential for engulfment, as many dying cells are still properly removed in these mutants. Genetic analysis of various double mutants has suggested that these six genes might form two partially redundant groups, one being comprised of CED-1, CED-6, and CED-7; the other of CED-2, CED-5, and CED10 (Ellis et al., 1991). The number of persistent cell corpses is increased dramatically in double mutants crossing groups, but not in those within the same group. Understanding how these genes are involved in regulating engulfment requires the elucidation of their molecular nature. In other species, several candidate apoptotic receptors have been identified over the past few years; these include the ATP-binding cassette transporter ABC1 (Luciani and Chimini, (1996), EMBO J. 15 pp 226-235) adhesion molecules such as the vitronectin receptor (Savill et al (1990), Nature 343 pp 170-173) and CD36 (Asch et al. (1987) J. Clin. Invest. 79 pp 1054-1061; Savill et al (1992) J. Clin. Invest. 90 pp 1513-1522; Ren et al (1995) J. Expl Med. 18 1857-1862), Drosophila croquemort (Franc et al., (1996), Immunity 4, pp 431-443 class A scavenger receptors (Platt et al., (1996), Proc. Natl. Acad. Sci. USA 93 pp 12456-12460) lectins (Duvall et al., (1985), and a predicted receptor that can recognize phosphatidylserine on the outer leaflet of apoptotic cells (Fadok et al., (1992) J. Immunol. 148 pp 2207-2216; Fadok et al (1992) J. Immunol 149 pp 4029-4035). Currently little is known about the molecules used by engulfing cells to transduce signals from surface receptors to the cytoskeleton, or how these molecules regulate the local cytoplasmic rearrangements and dynamic extensions that are required for phagocytosis (Savill et al., 1993). A genetic analysis of engulfment in C. elegans could identify genes involved in these processes. Indeed Wu and Horvitz (1998) (Nature 3.92 pp 501-504) showed that C. elegans CED-5 is homologous to human DOCK180, and might regulate cytoskeleton rearrangement during engulfment. The process of apoptosis has been implicated in the etiology, or associated with the pathology, of a wide range of diseases, including cancer, autoimmune diseases, various neurodegenerative diseases such as Amyotrophic Lateral Sclerosis, Huntington's Disease, and Alzheimer's Disease, stroke, myocardial heart infarct, and AIDS (Thompson, (1995) Science 267 pp 1456-1462). Thus, a better understanding of the molecular events that underlie apoptosis might lead to novel therapeutic interventions. While much of the current attention is centered on the genes and proteins that control the killing step of the death process, it is very likely that the removal of apoptotic cells will prove to also be crucial for the proper overall functioning of the apoptotic program, and will offer another entry point for therapeutic intervention (as described herein). The process of recognition and engulfment of dying cells is extremely swift and efficient. In animals, it is essentially impossible to find a cell with apoptotic features that is not already within another cell. Such rapid recognition and phagocytosis of apoptotic cells is a crucial aspect of programmed cell death in vivo: unengulfed apoptotic bodies can undergo secondary necrosis, leading to inflammation. Failure to remove apoptotic bodies also exposes the body to novel epitopes (from e.g., caspase-generated protein fragments), possibly encouraging the development of autoimmune disease. Persistent apoptotic bodies can often be observed following chemotherapeutic intervention (which leads to extensive apoptosis) and are particularly abundant in solid tumors, in which clearance of cell corpses might be delayed. In addition to their ability to recognize and engulf apoptotic cells, professional phagocytes carry specific surface receptors, such as the Fc (Ravetch, (1994) Cell 788 553-560; Greenberg et al., (1993) J. Exp. Med. 177 pp 529-534) and C3 (Bianco et al., (1975) J. Exp. Med. 141 pp 1278-1290; Greenberg, (1995) Trends in Cell Biol. 5 pp 93-99) receptors, which recognize antigen-opsonized particles and trigger their phagocytosis. Inhibitor studies have shown that Fc receptor-mediated phagocytosis requires tyrosine phosphorylation (Greenberg et al., 1993; Greenberg, 1995). The work of the present inventors suggests that the engulfment of apoptotic cells could be also mediated by a tyrosine kinase signal transduction pathway. While these two pathways clearly use distinct receptors at the cell surface, they must eventually converge on the same downstream engulfment machinery, and could thus share at least some common signal transduction molecules. The invention relates to an isolated protein which is an adaptor molecule in a signal transduction pathway which regulates phagocytosis of apoptotic cells. In a particular embodiment, the invention pertains to an isolated protein from the nematode worm C. elegans which is an adaptor molecule acting in a signal transduction pathway which promotes phagocytosis of apoptotic cells, which protein comprises the amino acid sequence shown in Figure 2A (SEQ ID No: 2) or an amino acid sequence which differs from Figure 2A only in conservative amino acid changes. As aforesaid the amino acid sequence shown in Figure 2A is that of the C. elegans CED-6 protein with its encoding DNA also shown. In another of the aspects the invention comprises a nucleic acid comprising a sequence of nucleotides which encodes the amino acid sequence of Figure 2A, (SEQ ID No: 2) for example, a sequence of nucleotides from about nucleotide position 22 to about nucleotide position 1500 of Figure 2A or the entire sequence of nucleotides shown in Figure 2A. In a further embodiment of the invention there is provided an isolated protein which is a fragment or portion of a protein having the amino acid sequence of Figure 2A or of a protein having an amino acid sequence which differs from that shown in Figure 2A only in conservative amino acid changes. For example, the portion may comprise an amino acid sequence corresponding to the phosphotyrosine binding domain (SEQ ID No: 4) (about amino acid 46 to about amino acid 193 in Figure 2A) or an amino acid sequence corresponding to the proline/serine rich region (SEQ ID No: 6) (about amino acid 242 to about amino acid 339 in Figure 2A). Nucleic acids (SEQ ID Nos: 3 and 5 respectively) encoding the PTB domain or the proline/serine rich region of the C. elegans CED-6 protein are encompassed by the claimed invention. In yet a further aspect of the invention there is provided an isolated nucleic acid capable of hybridizing to the sequence of nucleotides of SEQ ID Nos: 1, 3, 5, 7, 9, 11, 13, 15 under conditions of low, medium or high stringency. It is to be understood that low stringency means approximately: 0.2 to 2xSSC; 0.1% SDS; 25° to50°C. In a further embodiment of the invention there is provided a fusion protein ,which Comprises as part of the fusion a protein having an amino sequence of SEQ ID No: 2, 4, 6, 8, 10, 12, 14, or 16 or an amino acid sequence which differs from the amino acid sequence shown in SEQ ID Nos: 2, 4, 6, 8, 10, 12, 14, or 16 only in conservative amino acid changes. The protein may be fused to, for example, an epitope tag or the expression product of a reporter gene. In yet a further aspect the invention provides expression vectors comprising any of the nucleic acid sequences ofSEQ ID Nos: 1, 3, 5, 7, 9, 11, 13, Preferably, the vectors incorporate a reporter gene such as green fluorescent protein which is positioned relative to the nucleic acid of the invention such that expression of the nucleic acid results in expression of the reporter gene. Preferably, a fusion of CED-6 and the reporter gene is expressed. It is to be understood that the term "nucleic acid" as used herein may include genomic DNA, RNA and cDNA. Positional cloning methods were used to clone the C. elegans CED-6 gene and determine the nucleotide sequence. In addition they have functionally characterized the protein. By searching publicly available protein sequence databases, it has been determined that the CED-6 protein has in the N-terminal half a putative phosphotyrosine binding domain and in the C-terminal half a proline/serine rich region which is a potential SH3 binding domain. Genetic mosaic analysis, as well as rescue and over-expression experiments, have shown that CED-6 acts autonomously within engulfing cells and promotes engulfment of apoptotic cells. Further database searching has confm'ned the functional regions to be surprisingly evolutionally conserved. Thus, the inventors have now cloned two human homologues of the C eiegans CED-6 gene and shown them to have equivalent function. Molecular Cloning of C. elegans CED-6 Previous genetic mapping experiments by Ellis and Colleagues (Ellis et al, (1991) (Genetics 129 pp 79-94) have placed CED-6 gene close to the daf-4 locus on chromosome three (Figure 1A). The region around daf-4 has been mostly sequenced by the C. elegans genome sequence consortium (Wilson et al, (1994) Nature 368 pp 32-38). To determine the exact physical location of CED-6, the present inventors collected thirteen overlapping cosmids in this region which together are roughly 0.3 Mbp. Using the germline transformation method (Mello and Fire, (1995), methods in cell biology (San Diego Academic Press) pp 452482)these cosmids were tested for their ability to rescue the engulfment defect of CED-6(n1813), by scoring three-fold embryos laid by transgenic animals for the presence of persistent cell corpses. Three fold embryos were chosen for the initial study because cell corpses are numerous and easily seen at this stage of development. Two overlapping cosmids F56D2 and F43F12 were found to be able to rescue the engulfment defect of CED-6(n1813). The further rescuing experiments using the DNA fragments from F56D2 were identified to contain the rescuing activity. The gene prediction program GENEFINDER TM suggested that this region contains two genes, which the C. elegans genome sequence consortium submitted to Genbank under the names F56D2.7 and C05D2.6. Using a combination of RTPCR and screening of cDNA libraries (see below) the existence and predicted intron/exon pattern of F56D2.7 was confirmed. However, the inventors found that C05D2.6, rather than corresponding to a single gene, actually corresponds to two genes and the short distance (>>>bp) between the end of the upstream transcript and the start of the downstream transcript suggested that C05D2.6A/B might be a two-gene operon (Zorio et al (1994) Nature 372 pp 270-272.). It was found that C05D2.6B is trans-spliced to the "downstream" splice leader SL2, whereas the upstream transcript C05D2.6A is trans-spliced to the more common SL1 splice leader (Figure 1E). The CED-6 Locus To determine which one of the three genes present on the Xho I fragment corresponds to CED-6, a number of constructs were generated containing internal deletions or point mutations. The deletion of most of the C05D2.6A/B operon had no deleterious effect on CED-6 rescue, whereas the introduction of a frameshift mutation within exon 3 of F56D2.7 abolished the fragment's rescuing activity (Figure 1E). To exclude the possibility that F56D2.7 might be a multicopy suppressor of CED-6, and to confirm suspicions that F56D2.7 might correspond to the CED-6 locus, the two known CED-6 alleles, n1813 and n2095 were analysed for any nucleotide changes within this region. Southern blot analysis revealed an allele-specific restriction fragment length polymorphism affecting F56D2.7 in CED-6(n2095) mutants (Figure 2A). Based on the hybridization patterns observed in n2095, a single nucleotide deletion in exon 4 of F56D2.7 in CED-6(n1813) was also identified. This mutation should result in a reading frame shift and a truncated protein (Figure 2B). Taken together, the genomic rescue and mutation data strongly suggested that F56D2.7 corresponded to CED-6. Identification of CED-6 Transcripts To confirm the predicted intron/exon structure for CED-6, the present inventor screened a mixed-stage cDNA library and identified 10 clones corresponding the CED-6 gene. Several of these contained splice leader SL2 sequences at the 5' end, suggesting that CED-6 might also be a downstream gene in an operon. RT-PCR was performed on mixed-stage RNA using both SL1 and SL2 trans-splicing leaders as primers for the PCR step. Interestingly, sequence analysis of the PCR-amplified fragments revealed that both SL 1 and SL2 transsplicing leaders can be found at the 5' end of CED-6 transcripts (Figure 2B). The upstream gene in the CED-6 operon is the predicted gene F56D2.1. The presence of SLl-trans-spiced mRNA suggests that CED-6 might also be transcribed from a second downstream promoter, independently of the upstream gene. The existence of a downstream promoter could explain why the Xho I fragment could rescue CED-6 mutants even though it does not contain the whole CED-6 operon. CED-6 Protein Contains a Phosphotyrosine Binding (PTB) Domain and a Proline/Serme Rich Region The full-length CED-6 cDNA is predicted to code for a 492 amino acid protein (Figure 2B). A search of public sequence database with the predicted CED-6 sequence indicated that the N-terminal half of CED-6 contains a putative phospho-tyrosine binding (PTB) domain. PTB domains can promote binding to phosphorylated tyrosine residues located within an appropriate primary sequence context. The PTB domain is similar in function, but distinct in structure from the SH2 domain. The present inventors have aligned the CED-6 PTB domain with the PTB domains found in a number of other proteins (Figure 3A). Secondary structure prediction programs suggest that most of these structural elements also exist in the CED-6 PTB domain. In addition to its similarity to known proteins, the CED-6 PTB domain also showed significant sequence similarity to the predicted translation products of a number of expressed sequence tags (ESTs; Figure 3A, B). In fact, the degree of similarity between CED-6 and a number of these ESTs was much higher than between CED-6 and any previously characterized protein (Figure 3A, 3B). Furthermore, in several cases, the sequence similarity between CED-6 and ESTs extended beyond the PTB domain (Figure 3B). CED-6 also contains a proline/serine rich region at its C-terminal half, with 42°,/o serine over a 24 amino acids stretch and clusters of proline-rich regions (Figure 2B, Figure 3B). These proline-rich regions were characterized by several sequence signatures of PxxP (Figure 2A), which has been shown to promote interaction with SH3 domains (Ren et al, (1993); Yu et al (1994) Cell 76 pp 933-945,; Grabs et al (1997) J. Biol, Chem. 272 pp 13419-13425). Between the PTB and proline-rich regions is a short stretch rich in charged residues(41% charged amino acids over 46 amino acids). This highly charged region is also found in several other PTB domain containing proteins, including mouse p96, Shc, and C. elegans M110.5 (Figure 3B). Conservation of CED-6 Amongst Species It was found that these EST clones also shared the homology region beyond the PTB domain with the CED-6 protein. A C. Briggsae EST clone has 72% identity to CED-6 over 132 amino acids at the N-terminus, and 64% identity to CED-6 over 103 amino acids at the C-terminus (Figure 3B). Three overlapping human EST clones were also obtained and constructed into one sequence. The human EST fusion sequence showed -54% identity to PTB domain of CED-6, and also contains a highly charged region right after the PTB domain. The evolution tree based on the alignment of PTB domains showed that CED-6 formed a subgroup with EST clones from human, Drosophila, and C. Briggsae, suggesting that these proteins might be functionally conserved. Mouse p96, Drosophila Disabled, and C. elegans M110.5 formed another subgroup (Figure 3C). The tree also indicated that the Shc subgroup is more similar than the p96 subgroup to CED-6 subgroup. CED-6 Acts Cell-autonomously Within Engulfing Cells A genetic mosaic analysis was performed to determine if CED-6 acts within engulfing cells or dying cells. For convenience, a pair of cells on adult gonad, germ cells and somatic sheath cells (Figure 4A) were used. During oogenesis large number of oocytes undergo programmed cell death, and normally these dying cells are engulfed by somatic sheath cells (Hengartner,1997). In this analysis a mosaic pattern of genetic background for CED-6 and wild type between germ cells and somatic sheath cells was generated. Ncl-1 mutant was used for the identification of the mosaic pattern in the single-cell resolution since in the Ncl-1 mutant somatic cells of animals exhibit abnormal enlarged nucleoli, which can be easily identified under Normaski optics (Herman, 1984; Genetics 108 pp 165-189; Hedgecock and Herman, 1995 Genetics 141 pp 989-1006). A strain was constructed dpy-17(e164) CED-6 (n 1813) mec14(u55) ncl-1(e1865) unc-36(e251)IlI; sDp3. This worm strain showed a wild type phenotype since the sDp3(l11;.D duplication covers all these mutations (Rosenbluth et al, (1985) Genetics 109 pp 493-511). To identify the animals with CED-6 mutant germ cells and wild-type somatic sheath cells, animals must be found with the duplication loss from any of P2, P3 and P4 lineages but not from EMS, MS or any lineages below the MS which would lead to the loss of the duplication in somatic sheath cells (Figure 4). These animals can be obtained by looking through many animals of the constructed strain for the animals laying only Dpy Unc progenies. The animals with the loss of the duplication in P 1 lineage also lay only the Dpy Unc progenies, however these animals are not mosaic animals for the present purpose since the loss of the duplication in P1 lineage results in the CED-6 mutant background in both germ ceils and somatic sheath cells. From 1,000 dpy-17(e164) CED-6(n1813) mec-14(u55) ncl-1(e1865) unc-36(e251)111; sDp3 animals, six animals were identified laying only Dpy Unc progenies. Observation of these six animals under Normaski optics indicated that one animal had the duplication lost in P4, one in P3, three in P2, and one in P1. All five animals displayed no cell corpses in gonad except the one with the duplication lost in P1, suggesting that CED-6 is not required in germline for engulfment. Since the chance for loss of the duplication in all cell divisions is approximately the same (Hedgecock and Herman, 1995), the rate of the sDp3 loss is 0.15% per cell division. Animals were then looked for with the CED-6 mutant somatic sheath cells and wild-type germ cells. From 500 animals four animals were identified with enlarged nucleoli in the somatic sheath cells in one arm of the gonad (Figure 5B), and all four animals did not have the duplication lost in the lineage generating germ cells (Figure 4). Three animals appeared to have the duplication lost in sheath cells in the anterior arm but not in the posterior arm. And the accumulated cell corpses were only observed within the anterior gonad arm, but not the posterior gonad arm of these animals (Figure 4, Table). One animal had the duplication lost in the sheath cells surrounding the posterior gonad arm, but not in that surrounding the anterior arm. This animal had cell corpses accumulated within the posterior arm but not the anterior arm (Figure 4). These results suggest that CED-6 is required for somatic sheath cells, or engulfing cells to eliminate the dying cells in adult gonad. CED-6 Promotes the Engulfment of Embryonic and Germ Cell Corpses To unambiguously demonstrate that F56D2.7 eDNA indeed corresponds to CED-6, the inventors tested whether the full-length F56D2.7 eDNA can rescue the engulfment defect of CED-6 mutants, and transgenic animals were generated carrying the F56D2.7 cDNA under the control of the C. elegans heat shock promoters hsp-16.2 and hsp-16.48 (see Examples) Used together, these two promoters drive expression in almost all somatic cells, including both cells that normally undergo programmed cell death and cells that normally engulf the dying cells. To test for rescue, embryos laid by transgenic mothers were exposed to a brief heat shock pulse just prior to the appearance of the first developmental cell deaths, and scored the number of persistent corpses visible in the heat-shocked animals after hatching (Figure 4). As expected, over-expression of F56D2.7 eDNA significantly and specifically reduced the number of persistent cell corpses visible in CED-6 mutants, confirming that F56D2.7 is the relevant gene affected by the mutations that we detected in CED-6(n 1813) and CED-6(n2095) mutants. Rescue of F56D2.7 eDNA in germline was also tested (Figure 5C). Adult hermaphrodites were exposed to a brief heat shock pulse just prior to the appearance of the germline cell death, and scored the number of persistent cell corpses 12 hours and beyond after the heat shock. No cell corpses were found in gonads of the majority of animals, suggesting that CED-6 eDNA can also rescue the engulfment defect of CED-6 in germline. Recognition and engulfment of apoptotic cells is a very early event in C. elegans programmed cell death (Robertson and Thomson, (1982)J. Embryol. Ex. Morph 67 pp 89-100). In CED-6 mutants, the extension of cytoplasm is blocked, resulting in the persistence of cell corpses (Ellis et al, 1991). These cell corpses, however disappeared from the animal eventually. To determine whether CED-6 acts only in a narrow time-window at the early stage of cell death or whether the signal transduction pathway can be used to engulf cell corpses formed many hours after cell death takes place, the inventors tested whether F56D2.7 cDNA promotes the engulfment of persistent cell corpses. CED-6 was over-expressed three hours before the embryos hatch, when most of cells dying by programmed cell death during the embryonic development have been dead approximately for five hours (Figure 5A), and examined cell corpses three laours after the heat-shock on the head of L1 larvae. The number of cell corpses was found to be suppressed significantly (Figure 5B). The control experiments with either no heat treatment, or over-expression of lacZ showed no obvious effect on the corpse expression, suggesting that over-expression of CED-6 can promote the engulfment of cell corpses in soma (Figure 5B). The inventors also tested if over-expression of CED-6 could promote the engulfment of cell corpses formed hours after the cell death in the germline (Figure 5D). Adult transgenic animals carrying CED-6 cDNA driven by the heat shock promoters were heat structured at several time points after the accumulation of cell corpses in gonad and the number of cell corpses 12 hours after the heat shock were examined. It was found that cell corpses could be removed sufficiently at all time points, suggesting that over-expression of CED-6 can promote the engulfment of cell corpses accumulated in germline for hours, even days (Figure 5D). The present inventors have concluded that the signal transduction pathway in which CED-6 is involved can carry on the task of removing cell corpses, and there is no specific time-window for CED-6 to act during the process of programmed cell death. Mosaic CED-6 Protein Expression Supports That CED-6 Acts Within Engulfing Cells The invention includes methods to detect quickly if CED-6 acts within engulfing cells. This method is based on dying cells' failing to express proteins so as to generate a mosaic pattern of protein expression. However, this idea can be only applied to the soma, but might not to the germline, since in germline all germ cells share one syncytial cytoplasm (Hirsh et al, (1976) Developmental Biology 49 pp 200-210), so those germ cells carrying the transgenes could contribute the expressed proteins into the cytoplasm, subsequently all newly formed oocytes. However the mosaic pattern of the protein expression can be generated in the germline because the transgenes have been found not to be expressed well in germ cells. The expression pattern of heat shock promoters in gonad were examined. Adult animals carrying the lacZ transgenes driven by heat shock promoter were applied heat shock 24 hours after L4 molts. The lacZ expression by beta-gal staining in both germ cells and sheath cells was subsquently examined. It was found that somatic sheath cells were stained blue and the stain could last 60 hours after the heat shock, but not the germline at any time point after the heat shock, the similar result was also observed in previous studies (Stringham et al, (1992) Molecular Biology of the cell 3 221-233). The expression of CED-6 in germline upon heat shock was also examined for three-fold embryos laid by heat-treated transgenic animals for the rescuing activity of the engulfment defect. It was found that the majority of embryos had the CED-6 mutant phenotype, suggesting that CED-6 is not expressed well in germline. That CED-6 transgene in gonad is not expressed very well provided a useful tool to test if CED-6 acts within the somatic sheath cells. As described in Figures 4 and 5, cell corpses were not observed in majority of animals in gonad at the different time point after the heat treatment, and the phenomenon lasted until 60 hours or beyond after the heat treatment (5C). In contrast to this result, without the heat treatment these transgenic animals had cell corpses accumulated in gonad, similar to that of the CED-6(n1813) mutant. Over-expression of lacZ didn't affect the expression of cell corpses of CED-6 mutant, either (5C). These results support the conclusion from the mosaic analysis that CED-6 might act within engulfing cells, the somatic sheath cells. This method provides a simple way to detect if a gene acts within engulfing cells or dying cells. Site of active of CED-6 in relation to CED-1 and CED-7 To understand if CED-6 genetically interacts with any other engulfment genes, CED-6 was over-espressed at the genetic background of CED-1, 7, 2, 5, and 10. The extra-chromosomal arrays carrying CED-6 cDNA driven by heat shock promoters were transferred from CED-6(nl813) background to wild-type N2 background, and subsequently to CED-1, 7, 2, 5, and 10 mutant background. CED-6 was then over-exposed by following the method used for the rescue of CED-6 engulfment defect by the over-expression of CED-6 cDNA as described Figure 5A. It was found that over-expression of CED-6 could partially suppress the engulfment defect for CED-7(n1997). To understand if the suppression is allele-specific, two additional alleles, CED-7(n1996) and CED-7(n1892), were tested and similar results were achieved, suggesting that the suppression is not allele-specific (Figure 6). For the same purpose three alleles of CED-1, n1506, n1995, and n1735, were also tested it was found that over-expression of CED-6 could partially suppress the engulfment defect of three alleles of CED-1 (Figure Several control experiments were performed to confirm that these rescue were specific for CED-6. Transgenic animals with CED-6 transgene without heat treatment were tested; over-expression of lacZ at CED-1 or CED-7 engulfment mutant background was also tested. Results showed that the similar numbers of cell corpses were achieved as that of the CED-1 or CED-7 mutants. Heat treatment reduced the expression of cell corpses for CED-7(n1997). Overexpression of CED-6 reduced the expression of cell corpses even more. These data suggest that the partial suppression of the engulfment defect of both CED-1 and CED-7 are specific for CED-6. It was also observed that over-expression of CED-6 did not have obvious effect on the number of cell corpses for CED-2, 5 and 10. These results suggested that CED-6 might act downstream of both CED-1 and CED-7, and CED-2, 5 and 10 act either downstream of CED-1, 6, and 7 or in a different pathway (Figure 6). The Regulation of the CED-6 Expression SL2 was detected at the 5' end of the CED-6 cDNA, suggesting that CED-6 is a downstream gene of an operon (Huang and Hirsh, (1989); Proc. Natl, Acad. ScI.USA 86 pp 8640-8644; Spieth et al (1993) Cell 73 pp 521-532; Zorio et al (1994) Nature 372 pp 270-272; Blumenthal et al (1995) TIG II pp 132-136). The inventors have shown previously that a 10 kb Xho I fragment can rescue the engulfment defect of the CED-6 mutant. The fragment, however contains only CED-6, the downstream gene of an operon, but not the upstream one. The expression of CED-6 might rely on the 1 kb upstream region of CED-6 gene, a intergenic region of the operon. The Intergenic region of a operon sometimes could be used as a promoter for the expression of the downstream gene (Blementhal and Steward, (1997 C.elegans II) (Cold Spring Harbor; Cold Spring Harbor Laboratory Press pp 117-145) CED-6 is an Adaptor Molecule Acting in the Signal Transduction Pathway of the Engulfment Protein phosphorylation is a well-defined "switch" mechanism for cells to deliver signals from one protein to another, and it is essential to transduce extracellular signals inside cells. PTB domain is another domain besides the SH2 domain to be able to interact with a phosphorylated tyrosine residue (Kavanaugh and Williams, (1994) Science 266; Blaikie et al, (1994) J.Biol.Chem 269 3203132034). Several proteins containing PTB domains have been found to act as adaptor molecules in the signal transduction pathway. These include Shc, Sck, Numb, FE65, disabled, DOC-2, P96 and IRS-1 (Bork and Margolis, (1995) Cell pp 693-694); Geer and Pawson, (1995) TIBS 20 pp 277-280). The proline rich region from many proteins have been shown to form multiproline helix and interact with a SH3 domain (Ren et al, 1993; Gout et al, (1993) Cell 75 pp 25-36; Yu et al, 1994). Both biological analysis and analysis of the crystal structure of the SH3 binding domain suggested that the sequence signature, PxxP, was essential for its interaction with the SH3 domain (Ren et al, 1993; Yu et al, 1994; Grabs et al, 1997). CED-6 contained stretches of proline rich regions containing the PxxP signature, suggesting its potential to interact with the SH3 domain. CED-6 is an adaptor molecule that directly or indirectly transduces the signal from receptors to effectors or cytoskeleton molecules to initiate the engulfment process. The Interaction Partners of CED-6 The PTB domain has been shown to interact specifically with a NPXY(p) motif (Kavanaugh and Williams, 1994; Zhou et al, (1995) Nature 378 pp 584-592; Geer and Pawson, 1995). Many receptors such as EGF receptor, TrkA, insulin receptor, IGF-1 receptor contain this motif at the carboxyl terminal (Geer and Pawson, 1995). Signals from these receptors have been shown to be transduced through the interaction of a phosphotyrosine residue of this motif with PTB domains of adaptor molecules, such as Shc and insulin receptor substrate 1. The inventors found that in the intracellular region of CED-7 there was a NPXY(p) motif. CED-7 has been suggested to act in the same genetic pathway with CED-6 (Ellis et al, 1991). The inventors have shown that CED-7 might act upstream of CED-6 (Figure 7). CED-7 encodes a ABC transporter, and its mammalian homologue, ABC1 was found to be required for the macrophage to engulf dying cells (Luciani and Chimini, 1996), suggesting that CED-7 might act within engulfing cells. It is possible for CED-6 to physically interact with CED-7 through a PTB domain with NPXY(p) motif of CED-7 to regulate the signal transduction of engulfment process. CED-6 also contains a proline/serine rich region with several sequence signature PxxP, which might mediate its interaction with the SH3 domain. The SH3 domain has been suggested to mediate protein-protein interactions between signaling molecules downstream of membrane-bound receptors (Koch et al, (1991) Science 252 pp 252-673; Pawson and Schlessinger, (1993) Current Biology 3 pp 434-442. A SH3 domain containing protein is likely to interact with CED-6 and to regulate the signal transduction pathway of engulfment. Several proteins might directly or indirectly interact with CED-6 protein. CED-1 might act upstream of CED-6(Figuie 6 & 7A). The relationship between CED-1 and CED-6 will depend on the cloning of the gene. A protein with a phosphorylated tyrosine residue should exist to interact with the PTB domain of CED-6. This phosphorylated protein is either a tyrosine kinase or a substrate of a tyrosine kinase, and a tyrosine phosphatase should also be involved in the signal transduction pathway of engulfment to down-regulate the activity of the phosphorylated proteins. Some studies on phagocytosis in mammalian system have shown that a tyrosine kinase signal transduction pathway might play an essential role in the opsonin-mediated phagocytosis process (Roshenshine and Finlay, (1993) BioEssays 15 pp 17-24; Greenberg, (1995) Trends in Cell Biology 5 pp 93-99. The present results suggest that it might be the same case for the PCD triggered engulfment. These two types of phagocytosis might share some similarity at the end. CED-6 Acts Within Engulfing Cells A genetic mosaic analysis has been perfomaed to determine that CED-6 acts within engulfing cells. This conclusion was drawn based on the observation of a pair of cells, germ cells and somatic sheath cells. We have shown previously that over-expression of CED-6 can promote the engulfment of cell corpses. Since cells that have been dead for many hours are very unlikely to maintain their ability for protein expression (Estus, 1994; Freeman, 1994), the rescue of cell corpses is most likely to be due to the expression of CED-6 within the engulfing cells. This result suggests that CED-6 also acts within the engulfing cells in the soma. Previously it has been shown by the inventors that over-expression of CED-6 could rescue the engulfment defect of CED-6 in both soma and germline (Figure 5), suggesting that CED-6 acts in a similar mechanism in both places. CED-6 Can Promote the Engulfment of Cell Corpses Over-expression of CED-6 promotes the engulfment of dying cells at a very early stage of the cell death, and cell corpses formed hours after the cell death. Cell corpses have been shown to have a typical morphology ofapoptotic cells, for instance, membrane blebing. The antigens presented on the membrane surface of cell corpses for their recognition by engulfing cells might be somewhat different from that on the membrane surface of the early dying ceils. Irrespective of ligands on dying cells and receptors on the engulfing cells are the same or not in both situations, CED-6 is required for the engulfment. A few cell corpses in the gonad were not removed upon heat shock for some animals later after the heat shock. These corpses tend to be located in between oocytes and closed to the spermatheca. The failure of the engulfment of these cell corpses might be due to their lack of contact with the sheath cells. It is concluded that cell corpses, just like dying cells at the early stage of the PCD, can trigger phagocytosis. In mec-4 mutant animals six touch sensory neurons die of necrotic death due to a channel defect leading to an impaired osmotic pressure in these cells (Driscoll and Chalfie, (1991) Nature 349 pp 588-593). Chung and Driscoll showed that the removal of the swelling dead cells was delayed significantly at the CED-6 background, implying that CED-6 is also involved in the removal of necrotic dying cells. Thus, there might be similar signals presented on the surface of dead cells to allow them to be recognized by engulfing cells regardless the manner of the death; and the signal transduction pathway in which CED-6 is involved can be used to respond to these signals to cause engulfment. The fact that engulfment is triggered so early and is completed so swiftly is a clever design of nature, it is important especially for tissues with massive cell death. Conservation of the Engulfment Program In an alignment, an EST clone from C. Briggsae is highly conserved with CED-6 in both the Nand C-terminal region, suggesting that this EST clone might represent a real CED-6 homologue (Figure 3B). EST clones for Drosophila and human are also highly conserved to CED-6 but mainly in the region of PTB domain (Figure 3A & 3B). This result suggested the possibility for these PTB domain proteins to be functional homologues of CED-6 in those specimens. As a result two human homologues of C.elegans CED-6 gene have been cloned and characterized. Expression Vectors and Transfected Mammalian Cells Expressing CED-6 Fragments of C.elegans CED-6 DNA was inserted into commercially available vectors, including vectors having the reporter gene, green fluorescent protein (GFP), are set out in table 1 below; TABLE 1 GFP-CED-6 expression in MCF7 Cloning of CED-6 fragments in pEGFP from...(bp)-to .... (b-) Vector 2-1591 22-1492 598-1581 598-1494 22-745 744-1581 744-1494 TA-PCR pGA 1 pGA2 pGA3 pGA4 pGA5 pGA6 pGA7 pAS2 pGAI011 pGA1013 pGA D414 pEGFP-C 1 (*) pGA3011 pGA3013 pGA3015 pGA3036 pEGFP-C3(*) pGA3045 pEGFP-N3(*) pGA3062 pGA3064 pGA3067 pEGFP-N3(*) *are commercially available from Clontech Visualization GFP fluorescence in MCF7 cells Human breast cancer cells, MCF7 (ATCC: HTB-22), were seeded in Lab Tek chambered coverglass (Nalge Nunc International) and transfected using lipofectAMINE (GibcoBRL). After 18 hours, the chambered coverglasses where placed on a inverted microscope, and GFP fluorescence could be visualized. Expression of GFP-CED-6 Subcellular localization of worm CED-6 was assayed using GFP fusion proteins. By using different fragments the inventors showed that CED-6 has a clear cytoplasmic localization. This localization was abolished when only the PTB of CED-6 was used indicating that the C-terminal part might be implicated in proper targeting. Since the actual expression level varies from cell to cell one can observe an apoptotic phenotype in highly expressing cells and an elevated level of phagocytosis in strong expressing cells. In addition, localization to the lamelli was observed in some cells which perform engulfment. The transfected MCF7 cells as above are useful for conducting assays to identify compounds which inhibit and enhance CED-6 or CED-6 as will be discussed hereafter. Human Homologues of C. Elegans CED-6 In accordance with the invention there is provided an isolated protein which is an adaptor molecule in a signal transduction pathway which regulates phagocytosis of apoptotic cells. In accordance with another embodiment of the invention there is provided an isolated protein which is a human homologue of C.elegans CED-6 which comprises an amino acid sequence as shown in Figure 20 or Figure 22 (SEQ ID No: 8) or an amino acid sequence which differs from that shown in Figure 20 only in conservative amino acid changes (hlCED-6). Also provided is a nucleic acid (DNA RNA, cDNA or genomic DNA; SEQ ID NO: 7, 13, 15) encoding hlCED-6, h2CED-6 or h3CED-6 (SEQ ID Nos: 8, 14, 16) or a functional equivalent thereof. For example the invention encompasses a nucleic acid comprising the sequence of nucleotides from about nucleotide position 430 to about nucleotide position 1344 shown in Figure 18, Figure 19, or Figure 22 or the entire sequence of nucleotides shown in these figures. The invention includes the open reading frame of the nucleic acid sequence that encodes c. elegans CED-6, CED-6, h2CED-6 or h3CED-6. The invention also provides a protein which is a fragment of the protein with the amino acid sequence shown in Figure 20, Figure 22 or Figure 29 (SEQ ID No: 8, 14, 16). The fragment may comprise a sequence of amino acids corresponding to the phosphotyrosine binding domain ofSEQ ID NOs: 8, 14, 16. For example, the PTB domain ofSEQ ID Nos: 8 or 16 is from amino acids Nos. 15-157. The invention also pertains to the nucleic acid and amino acid sequences of the proline/serine rich domains ofhlCED-6 and/or h3CED-6 (e.g., amino acid Nos.: 201-276 in Figures 20, 22, or 29). Similarly, the highly charged region ofSEQ ID NOs.: 8 or 16 is encompassed by the invention (e.g., amino acid Nos. 161-195 of Figures 20, 22 and 29). The invention includes the nucleic acid sequences that encode these fragments. There is also identified herein a splice variant ofh3CED-6 (referred to herein as h2CED-6) which variant comprises an amino acid sequence as shown in Figure 21 (SEQ ID No: 14) or an amino acid sequence which differs from that shown in Figure 21 only in conservative amino acid changes. Also provided is a nucleic acid (DNA, RNA, cDNA or genomic DNA) encoding h2CED-6 (SEQ ID No: 13) or a functional equivalent thereof, for example a nucleic acid comprising from about nucleotide position 430 to about nucleotide position 1206 in Figure 19 or the entire nucleotide sequence shown in Figure 19. (SEQ ID No: 13) The human CED-6 amino acid sequence (SEQ ID NO: 16) is also shown in Figure 26. Amino acid sequence SEQ ID NO: 16 (human CED-6) and SEQ ID NO: 8 (hlCED-6) differ at amino acid No. 150. The nucleic acid sequence (SEQ ID NO: 15) that encodes human CED-6 is shown in Figure 30A-B. The claimed invention includes SEQ ID NOs: 15 and/or 16, the open reading frame ofSEQ ID NO.: 15, and the nuclic acid and amino acid sequence that encoded the functional fragments, (e.g., serine/protein rich region, the PTB domain or the highly charged domain), as described herein. The invention also provides a fusion protein in which one part of the fusion is a protein having an amino acid sequence as shown in any ofSEQ ID Nos: 8, 14 or 16 or a sequence differing from acid sequences only in conservative amino acid changes. The protein may be fused with, for example, an epitope tag or expression product of a reporter gene. 99/37770 PCT/US 9 9/01361 The present invention is intended to encompass CED-6 proteins (e.g., C. elegans CED-6, hl CED-6, h2 CED-6 and/or h3 CED-6) and polypeptides having amino acid sequences analogous to the amino acid sequences of CED-6. Such polypeptides are defined herein as CED-6 analogs (e.g., homologues), orthologs, or mutants or derivatives. Analogous amino acid sequences are defined herein to mean amino acid sequences with sufficient identity of CED-6 (e.g., C. elegans CED-6, hlCED-6, h2CED-6 or h3CED-6) amino acid sequence tO possess the biological activity of CED-6. For example, an analog polypeptide can be produced with "silent" changes in the amino acid sequence wherein one, or more, amino acid residues differ from the amino acid residues of the CED-6, yet still possesses the biological activity of CED-6. Examples of such differences include additions, deletions or substitutions of residues of the amino acid sequence of CED-6. Also encompassed by the present invention are analogous polypeptides that exhibit greater, or lesser, biological activity of the CED-6 proteins of the present invention. The claimed CED-6 protein and nucleic acid sequences include homologues, as defined herein. The homologous proteins and nucleic acid sequences can be determined using methods known to those of skill in the art. Initial homology searches can be performed at NCBI against the GenBank (release 87.0), EMBL (release 39.0), dbEST SwissProt (release 30.0) databases using the BLAST network service and other EST databases. Altshul, SF, et al, Basic Local Alignment Search Tool., J. Mol. Biol. 215:403 (1990), the teachings of which are incorporated herein by reference. Computer analysis of nucleotide sequences can be performed using the MOTIFS and the FindPatterns subroutines of the Genetics Computing Group (GCG, version 8.0) software. Protein and!or nucleotide comparisons can also be performed according to Higgins and Sharp (Higgins, D.G. and P.M. Sharp, "Description of the method used in CLUSTAL," Gene, 73:237-244 (1988)). Homologous proteins and/or nucleic acid sequences to the CED-6 protein and/or nucleic acid sequences that encode the CED-6 protein are defined as those molecules with greater than 70% sequences identity and/or similarity (e.g., 75%, 80%, 85%, The "biological activity" of CED-6 proteins is defined herein to mean the ability to regulate or affect the phagocytosis of apoptotic cells. The claimed CED-6 proteins also encompasses biologically active polypeptide fragments of the CED-6 proteins, described herein. Such fragments can include only a part of the full-length amino acid sequence of an CED-6 yet possess the ability to modulate or regulate phagocytosis of apoptotic cells. For example, polypeptide fragments comprising deletion mutants of the CED-6 proteins can be designed and expressed by well-known laboratory methods. Such polypeptide fragments can be evaluated for biological activity, as described herein. Antibodies can be raised to the CED-6 proteins and analogs, using techniques known to those of skill in the art. These antibodies polyclonal, monoclonal, chimeric, or fragments thereof, can be used to immunoaffinity purify or identify CED-6 proteins contained in a mixture of proteins, using techniques well known to those of skill in the art. These antibodies, or antibody fragments, can also be used to detect the presence of CED-6 proteins and homologs in other tissues using standard immunochemistry methods. In particular, biologically active derivatives or analogs of the above described proteins, including fragments and functional domains from c. elegans CED-6, hlCED-6, h2CED-6, or h3CED-6, referred to herein as peptide mimetics, can be designed and produced by techniques known to those of skill in the art. (see e.g., U.S. Patent Nos. 4,612,132; 5,643,873 and 5,654,276, the teachings of which are incorporated herein by reference). These mimetics can be based, for example, on a specific CED-6, hlCED-6 or h2CED-6 or h3CED-6 amino acid sequence and maintain the relative position in space of the corresponding amino acid sequence. These peptide mimetics possess biological activity similar to the biological activity of the corresponding peptide compound, but possess a "biological advantage" over the corresponding CED-6 amino acid sequence with respect to one, or more, of the following properties: solubility, stability and susceptibility to hydrolysis and proteolysis. Methods for preparing peptide mimetics include modifying the N-terminal amino group, the C terminal carboxyl group, and/or changing one or more of the amino linkages in the peptide to a non-amino linkage. Two or more such modifications can be coupled in one peptide mimetic molecule. Modifications of peptides to produce peptide mimetics are described in U.S. Patent Nos. 5,643,873 and 5,654,276, the teachings of which are incorporated herein by reference. Other forms of the hi, h2, or h3 CED-6 proteins, encompassed by the claimed invention, include those which are "functionally equivalent." This term, as used herein, refers to any nucleic acid sequence and its encoded amino acid which mimics the biological activity of the hi, h2, or h3 CED-6 proteins and/or functional domains thereof. Biologically active is used to describe a protein capable of regulating the phagocytosis of apoptotic cells. A polypeptide can be in the form of a conjugate or a fusion protein, both of which can be made by known methods. Fusion proteins can be manufactured according to known methods of recombinant DNA technology. For example, fusion proteins can be expressed from a nucleic acid molecule comprising sequences which code for a biologically active portion of the protein and its fusion partner, for example a portion of an immunoglobulin molecule. For example, some embodiments can be produced by the intersection of a nucleic acid encoding immunoglobulin sequences into a suitable expression vector, phage vector, or other commercially available vectors. The resulting construct can be introduced into a suitable host cell for expression. Upon expression, the fusion proteins can be isolated or purified from a cell by means of affinity matrix. Expression vectors incorporating any of the above mentioned nucleic acids including those designated SEQ ID Nos: 1, 3, 5, 7, 9, 11, 13 or 15, optionally with a reporter gene as aforesaid, are also provided by the invention. The present invention also encompasses isolated nucleic acid sequences encoding the CED-6 (e.g., C. elegans CED-6, hlCED-6, h2CED-6 or h3CED-6) proteins described herein, and fragments of nucleic acid sequences encoding biologically active CED-6 proteins. Fragments of the nucleic acid sequences, described herein, are useful as probes. Specifically provided for in the present invention are DNA/RNA sequences encoding CED-6 proteins, the fully complementary strands of these sequences, and allelic variations thereof. Also encompassed by the present invention are nucleic acid sequences, genomic DNA, cDNA, RNA or a combination thereof, which are substantially complementary to the DNA sequences encoding CED-6, and which specifically hybridize with the CED-6 DNA sequences under conditions of stringency known to those of skill in the art, those conditions being sufficient to identify DNA sequences with substantial nucleic acid identity. As defined herein, substantially complementary means that the sequence need not reflect the exact sequence of the CED-6 (e.g., C. elegans CED-6, hlCED-6, h2CED-6 or h3CED-6) DNA, but must be sufficiently similar in identity of sequence to hybridize with CED-6 DNA under stringent conditions. Conditions of stringency are described in e.g., Ausebel, F.M., et al., Current Protocols in Molecular Biology, (Current Protocols, 1994). For example, non-complementary bases can be interspersed in the sequence, or the sequences can be longer or shorter than CED-6 DNA, provided that the sequence has a sufficient number of bases complementary to CED-6 to hybridize therewith. Exemplary hybridization conditions are described herein. Cloning of human CED-6 Following the cloning of the C.elegans CED-6 gene and the full sequencing of the open reading frame, extensive searches against public domain human databases were performed. These revealed statistically significant homologies to a number of ESTs at the carboxy terminal region of the protein and one EST showed homology to the carboxy terminal of the PTB domain and at the beginning of the charged region. These ESTs were used for construction of primers for 5'RACE using a Marathonready cDNA colorectal adenocarcinoma library from Clontech. Subsequent additional sequence analysis and rounds of database searching revealed additional ESTs which enabled construction of a concensus sequence of approximately 2400 bp for h3CED-6 (Figure 6). Further sequence analysis has revealed a splice variant of the sequence shown in Figure 18 (h2CED-6), the portion which is alternatively spliced being underlined. The DNA of h2CED-6 is shown in Figure 19 and the amino acid sequence in Figure 21. The amino acid sequence of h2CED-6 is consistent with it being a dominant negative version ofhl or h3 CED-6 which antagonizes active of h l or h3CED-6. Assays for the identification of inhibitors and enhancers of CED-6 hlCED-6, h2CED-6, or h3CED-6 The cloning and functional characterization of C.elegans CED-6 and its two human homologues have permitted assay methods to be developed which allow identification of compounds which might inhibit or enhance CED-6, h 1CED-6, h2CED-6, or h3CED-6 activity or inhibit or enhance the transcription of these proteins. These may involve detection of the level of phagocytosis of apoptotic particles, measurement of level of actin-cytoskeleton rearrangement or detection of the level of transcription of the CED-6 proteins via a reporter gene such as GFP. An assay for the identification of inhibitors and/or enhancers of phagocytosis may consist of a cell line stably or transiently transfected with CED-6, hl CED-6, h2CED-6, or h3CED-6 or any other member of the CED-6 signal transduction pathway. Cell lines may also be microinjected with purified protein or vectors expressing antisense RNA. The expression product may be a fusion protein with GFP. Non transfected cells can be used in the assay also. The cell line may be a fibroblast cell line such as COSI, BHK 21, L929, CV1, Swiss 3T3, HT144, IMR32 or another fibroblast cell line. The cell line may also be an epithelial cell line such as HEPG2, MDCK, MCF7, 293, Hela, A549, SW48, G361, or any other epithelial cell line. The cell line may a primary line, such as human dermal FIBs, dermal keratinocytes, leucocytes, monocytes, macrophages, or any other primary cell line. Cells may be double transfected with other genes (like lectin, CD 14, SRA, CD36 ABC1, CED5, DOCK180) being from vertebrate (human fish, mouse) or invertebrate origin (C.elegans). Phagocytosis assays consist of the addition of and uptake of particles and/or apoptotic cells, by these cell lines. The particle may be opsonized heat or chemically killed bacteria and yeast in a variety of sizes, shapes and natural antigenicities. The particle or cell may be an opsonized, fluorescently labeled, heat or chemically killed bacteria and yeast in a variety of sizes, shapes and natural antigenicities. The cell may be a apoptotic neutrophils, apoptotic lymphocytes, apoptotic erythrocytes or any other apototic cell. These apoptotic cells may be opsonized and/or labeled with dyes or fluorescent dyes. The killed bacteria or yeast cells and the apoptotic cells are referred to as herein apoptotic particles. Assay 1 Cells, transfected with CED-6 or any other gene described herein, for example, nucleic acids ofSEQ ID Nos: 1, 3, 7, 9, 11, 13, or 15, can be grown in monolayer or in suspension. The apoptotic particles are added to the transfected cell. Phagocytosis can be followed by the uptake rate of the apoptotic particles. This can be measured by microscopy, by fluorescence microscopy, by quantitative spectrofluorometry and by flow cytometry. Cells and or particles may additionally be labeled with dyes, fluorescent dyes, antibodies and dyes of fluorescent dyes linked to antibodies prior to detection and measurement. Decrease or increase of the uptake of the apoptotic particles is a measurement for the influence of the transfected gene or genes in the phagocytosis. Assay 2 Compounds can be added to assay 1 to test their influence on the genes that are involved in the phagocytosis pathway. Transiently or stably transfected cells are grown in suspension or in monolayer. A series of compounds is added to the cells prior to the addition of the apoptotic particles. The influence of the compounds can be measured by comparing the uptake rate of the apoptotic particles with and without the addition of the compound. Measurements are described in Assay 1 Assay 3 Cells are able to phagocytose apoptotic particles by engulfment of particles. This involves the reorganization of the actin cytoskeleton. Mammalian cells, may be transiently or stably transfected with CED-6 or any gene involved in the CED-6 phagocytosis signal transduction pathway, for example, with a nucleic acid have the sequence ofnucleotides shown in any one of SEQ ID Nos: 1, 3, 5, 7, 9, 11, 13 or Cells can be any cell as described in Assay I. The genes may be expressed as a GPF fusion product. Cells may be double transfected (see Assay 1 ). The reorganization of the actin cytoskeleton can be visualized with fluorescent dyes linked to phalloidine, which interacts with F-actin. Reorganization of the cytoskeleton is an measurement for the engulfment induction by the transfected gene or genes. Transfected cells may be treated with particles or apoptotic cells as described in Assay 1. Reorganization of the cytoskeleton is visualized by microscopy or fluorescence microscopy. Assay 4 Compounds can be added to Assay 3 to test their influence on the genes that are involved in the cytoskeleton reorganization related to the phagocytosis pathway and engulfment. These compounds may enhance or inhibit the engulfment or cytoskeleton reorganization induced by the introduced genes. Transiently or stably transfected cells are grown in suspension or in monolayer. A series of compounds is added to the cells. The influence of the compounds can be measured by comparing the reorganization of actin cytoskeleton with and without the addition of the compound. Measurements as are described in Assay 1, Assay 2 and Assay 3. Apoptotic particles may be added in this test to induce phagocytosis, as described in Assay 2. Assay Non-transfected or transfected cell-lines such as those described above may be microinjected with purified CED-6 protein, for example, a protein having the amino acid sequence as shown in SEQ ID Nos: 2, 4, 6, 8, 10, 12, 14, or 16 or any protein from the CED-6 pathway or a fusion protein comprising any of said proteins. Microinjection can be done on the primary cell lines or the fibroblast cell lines or the other epithelial cells lines. The cell lines can be transfected with another gene prior to microinjections. Assays 1 through Assay 4 can be performed on these microinjected cells. Assay 6 Transfected or non-transfected cell-lines as described above may be micro-injected with a vector expressing CED-6 antisense RNA including antisense RNA in respect of any of the aforementioned proteins or any antisense RNA for genes involved in the CED-6 pathway. Microinjection can be done on the primary cell lines or the fibroblast cell lines or the epithelial cell lines. The cell lines can be transfected with another gene prior to micr0injection. Assays 1 through Assay 5 can be performed on these microinjected cells. Assay 7 Cell lines, as described in Assay 6 may be micro-injected with a vector expressing CED-6 antisense RNA or any antisense RNA for genes involved in the CED-6 pathway. Microinjection can be done on the macrophages. Inhibitory effects of the antisense RNA by inhibition of the CED-6 gene or genes involved in the CED-6 pathway can be followed and detected as described in Assay 1 through Assay 6. Compounds can be isolated which rescue the negative phenotype. Phagocytosis assays to screen for CED-6 inhibitor/enhancers in C.elegans The C.elegans CED-6 gene promotes the engulfment of dying embryonic and germ cells and persistent cell corpses. C.elegans may be used for detection and isolation of compounds that have an enhancing or inhibitory influence on phagocytosis and engulfment. In particular mutant worms lacking CED-6 activity or with otherwise altered CED-6 activity may be used or alternatively a transgenic worm transfected or transferred with CED-6, h 1CED-6, h2CED-6, or h3CED-6 DNA may be used. Assay 8 A series of compounds may be applied on CED-6 mutant worms or on worms harboring mutations in the CED-6 pathway. Restoration of engulfment induced by the compounds can be visualized using Nomarski microscopy by counting cell corpses remaining in the head region of L1 larvae and in the gonads of the worms. Assay 9 A series of compounds may be applied on humanized CED-6 mutant worms. Humanized worms are worms expressing the human CED-6 gene and are mutated for the C.elegans gene. Human CED-6 rescues the mutant phenotype. Compounds inhibiting or enhancing the CED-6 phenotype can be selected by visualization of the engulfment phenotype using Nomarski microscopy and looking for cell corpses as aforesaid. Medical applications The process of apoptosis has been implicated in the etiology - or associated with the pathology - of a wide range of diseases, including cancer, autoimmune diseases, various neurodegenerative diseases such as Amyotrophic Lateral Sclerosis, Huntington's Disease, and Alzheimer's Disease, stroke, myocardial heart infarct, and AIDS (Thompson, 1995). Thus a better understanding of the molecular events that underlie apoptosis might lead to novel therapeutic interventions. While much of the current attention is centered on the genes and proteins that control the killing step of the death process, it is very likely that the removal of apoptotic cells will prove to also be crucial for the proper overall functioning of the apoptotic program, and will offer another entry point for therapeutic intervention. The process of recognition and engulfinent of dying cells is extremely swift and efficient. In animals, it is essentially impossible to find a cell with apoptoic features that is not already within another cell. Such rapid recognition and phagocytosis of apoptotic cells is an crucial aspect of programmed cell death in vivo: unengulfed apoptotic bodies can undergo secondary necrosis, leading to inflammation. Failure to remove apoptotic bodies also exposes the body to novel epitopes (from e.g., caspase-generated protein fragments), possibly encouraging the development of autoimmune disease. Persistent apoptotic bodies can often be observed following chemotherapeutic intervention (which leads to extensive apoptosis) and are particularly abundant in solid tumors, in which clearance of cell corpses might be delayed. It is likely that failure to properly dispose of apoptoic cells leads to human disease. Genes involved in phagocytosis could therefore correspond to currently uncloned human inherited disease genes. Restoring proper phagocytosis would be a valid therapy for certain types of inflammation and autoimmune diseases. Conversely, In some cases, cells that should be maintained are inappropriately recognized by the engulfment machinery and cleared from the body. Preventing the engulfment of such cells could be of great therapeutic value. Examples of such diseases might include neurodegenerative diseases and stroke, as well as sickle cell aenemia. In addition activation of engulfment could be used for the same cases for which it is proposed to use activation of apoptosis, e.g., cancer. Indeed, specific activation within the cancer cells of the pro-engulfing signal would lead to the cells' removal - (and death) - without needing to activate the rest of the apoptotic machinery. This could be particularly useful for highly resistant tumors in which crucial elements of the central apoptotic machinery have already been inactivated. Thus, in accordance with another of its aspects the invention provides a method of treating, for example inflammation, autoimmune disease and cancer by administering to a patient an effective amount of a substance which enhances phagocytosis of apoptoic cells, in particular a substance which enhances the activity of h 1-CED6, h3-CED-6 or the signal transduction pathway in which it participates. Such substances includes hl-CED 6 or h3-CED-6 itself, a nucleic acid encoding hl- CED6 or h3-CED-6, an anti-sense nucleic acid to h 1, h2 or h3 CED-6 or compounds identified in any of the aforementioned assays as enhancers of CED-6, hl-CED-6, h2-CED-6, or h3-CED-6 or of transcription thereof. In addition the invention also enables a method of treatment of, for example, neurodegenerative diseases, stroke and sickle-cell anaemia by administering to a patient an effective amount of a substance which inhibits phagocytosis of apoptotic cells, in particular a substance which inhibits the activity ofhl-CED6 or h3-CED6 or the signal transduction pathway in which it participates. Such substances include h2 CED-6, a nucleic acid encoding h2CED-6, an anti-sense nucleic acid to hlCED-6 or h3CED-6 or compounds identified in any of the aforementioned assays as inhibitors of CED-6 or hl CED-6 or h3CED-6 or of transcription thereof. Pharmaceutical compositions comprising any of the above-mentioned therapeutic substances and a pharmaceutically acceptable cartier are also envisaged by the invention. -. To accomplish the various therapeutic treatments as described herein, a nucleic acid which encodes hi, h2 or h3 CED-6 or a functional portion or domain thereof must be introduced into a mammalian cell (e.g., mammalian somatic cell, mammalian germ line cell (sperm and egg cells)). This can be accomplished by inserting the isolated nucleic acid that encodes either the full length protein, or the domains described herein, or a functional equivalent thereof, into a nucleic acid vector, e.g., a DNA vector such as a plasmid, virus or other suitable replicon (e.g., a viral vector), which can be present in a single copy or multiple copies. The nucleic acid may be transfected or transformed into cells using suitable methods known in the art such as electroporation, microinjection, infection, and lipoinfection and direct uptake. Such methods are described in more detail, for example, in Sambrook et al., "Molecular Cloning: A Laboratory Manual," 2nd ED. (1989), Ausubel, F.M., et al., Current Protocols in Molecular Biology, (Current Protocol, 1994) and Sambrook et al., "Molecular Cloning: A Laboratory Manual," 2nd ED. (1989). hi, h2 or h3 CED-6 can be delivered to a cell by the use of viral vectors comprising one or more nucleic acid sequences encoding those proteins. Generally, the nucleic acid sequence has been incorporated into the genome of the viral vector. In vitro, the viral vector containing hi, h2 or h3 CED-6 protein described herein or nucleic acid sequences encoding the protein can be contacted with a cell and infectivity can occur. The cell can then be used experimentally to study phagocytosis of apoptotic cells or for assays as aforesaid or be implanted into a patient for therapeutic use. The cell can be migratory, such as hematopoietic cells, or non-migratory such as a solid tumor or fibroblast. The cell can be present in a biological sample obtained from the patient (e.g., blood, bone marrow) and used in the treatment of disease, or can be obtained from cell culture. After contact with the viral vector comprising the hl, h2 or h3 CED-6 protein or a nucleic acid sequence encoding them, the sample can be returned or readministered to a cell culture or patient according to methods known to those practiced in the art. In the case of delivery to a patient or experimental animal model (e.g., rat, mouse, monkey, chimpanzee), such a treatment procedure is sometimes referred to as ex vivo treatment or therapy. Frequently, the cell is targeted from the patient or animal and returned to the patient or animal once contacted with the viral vector comprising the activated mutant of the present invention. Ex vivo gene therapy has been described, for example, in Kasid, et al., Proc. Natl. Acad. Sci. USA 87:473 (1990); Rosenberg, et al., New Engl. J. Med. 323:570 (1990); Williams, et al., Nature 310476 (1984); Dick, et al., Cell 42:71 (1985); Keller, et al., Nature 318:149 (1985) and Anderson, et al., U.S. Patent No. 5,399,346 (1994). Where a cell is contacted In vitro, the cell incorporating the viral vector comprising a nucleic acid sequence of hl CED-6, h2 CED-6 or h3CED-6 can be implanted into a patient or experimental animal model for delivery or used in In vitro experimentation to study cellular events mediated by hi, h2 or h3 CED-6. Various viral vectors can be used to introduce the nucleic acid into mammalian cell. Viral vectors include retrovirus, adenovirus, parvovirus (e.g., adeno-associated viruses), coronavirus, negative strand RNA viruses such as orthomyxovirus (e.g., influenza virus), rhabdovirus (e.g., rabies and vesicular stomatitis virus), paramyxovirus (e.g. measles and Sendai), positive strand RNA viruses such as picornavirus and alphavirus, and double stranded DNA viruses including adenovirus, herpesvirus (e.g., Herpes Simplex virus types 1 and 2, Epstein-Barr virus, cytomegalovirus), and poxvirus (e.g., vaccinia, fowlpox and canarypox). Other viruses include Norwalk virus, togavirus, flavivirus, reoviruses, papovavirus, hepadnavirus, and hepatitis virus, for example. Examples of retroviruses include: avian leukosis-sarcoma, mammalian C-type, B-type viruses, Dtype viruses, HTLV-BLV group, lentivirus, spumavirus (Coffin, J.M., Retroviridae: The viruses and their replication, In Fundamental Virology, Third Edition, B.N. Fields, et al., Eds., Lippincott-Raven Publishers, Philadelphia, 1996). Other examples include murine leukemia viruses, murine sarcoma viruses, mouse mammary tumor virus, bovine leukemia virus, feline leukemia virus, feline sarcoma virus, avian leukemia virus, human T-cell leukemia virus, baboon endogenous virus, Gibbon ape leukemia virus, Mason Pfizer monkey virus, simian immunodeficiency virus, simian sarcoma virus, Rous sarcoma virus, lentiviruses and baculoviruses. A preferred method to introduce nucleic acid that encodes hl, h2 or h3 CED6 into cells is through the use of engineered viral vectors. These vectors provide a means to introduce nucleic acids into cycling and quiescent cells, and have been modified to reduce cytotoxicity and to improve genetic stability. The preparation and use of engineered Herpes simplex virus type 1 (D.M. Krisky, et al., Gene Therapy 4(10):1120-1125. (1997)), adenoviral (A. Amalfitanl, et al., Journal of Virology 72(2):926-933. (1998)), attenuated lentiviral (R. Zufferey, et al., Nature Biotechnology 15(9)871-875 (1997)) and adenoviral/retroviral chimeric (M. Feng, et al., Nature Biotechnology 15(9):866-870 (1997)) vectors are known to the skilled artisan. Hence, the claimed invention encompasses various therapeutic uses as aforesaid for the hi, h2 or h3 CED-6 protein or nucleic acid. The protein may be administered using methods known in the art. For example, the mode of administration is preferably at the location of the target cells. As such, the administration can be nasally (as in administering a vector expressing ADA) or by injection (as in administering a vector expressing a suicide gene tumor). Other modes of administration (parenteral, mucosal, systemic, implant, intraperitoneal, etc.) are generally known in the art. The agents can, preferably, be administered in a pharmaceutically acceptable carrier, such as saline, sterile water, Ringer's solution, and isotonic sodium chloride solution. The invention also provides diagnostic reagents which may be used in the diagnosis of a disease associated with a defect in phagocytosis of apoptotic cells. For example, an antibody to an epitope of any of the proteins with an amino acid sequence as shown in SEQ ID Nos: 2, 4, 6, 8, 10, 12, 14 or 16 could be used as a diagnostic reagent to determine whether a patient has a defect in h 1CED-6, h2CED6 or h3CED-6 or in the expression thereof. In addition defects at the genetic level can be detected by using as a probe a nucleic acid having a sequence as shown in SEQ ID Nos: 1, 3, 5, 7, 9, ll, 13, or 15 or portions thereof. Identification of the other proteins active in the CED-6 signal transduction pathway CED-6, hlCED-6, h2CED-6 or h3CED-6 can be used to identify other members of the signal transduction pathway promoting phagocytosis of apoptotic cells. There are number of possible methods by which this can be done but a preferred method is the so-called "two hybrid" system developed in yeast by Chien et al (1994, Proc. Natl. Acad Sci. USA 88 pp 9578-9582) which allows identification of proteins which bind to a particular protein of interest. This technique is based on functional in vivo reconstruction of a transcription factor which activates a reporter gene. More particularly the technique comprises providing an appropriate host cell, preferably yeast, with a DNA construct comprising a reporter gene under the control of a promoter regulated by a transcription factor having a DNA binding domain and an activating domain, expressing in the host cell a first hybrid DNA sequence encoding a first fusion of a fragment or all of a nucleic acid sequence according to the invention and either said DNA binding domain or the activating domain of the transcription factor, expressing in the host cell at least one second hybrid DNA sequence encoding putative binding proteins to be investigated together with the DNA binding domain or activating domain of the transcription factor which is not incorporated in the first fusion; detecting any binding of the protein being investigated with a protein according to the invention by detecting for the production of any reporter gene product in the host cell; optionally isolating second hybrid DNA sequence encoding the binding protein. EXAMPLES The N2 Bristol strain was used as the reference wild-type strain for this study. All strains were maintained as described by Brenner (Brenner, 1974), except that worms were raised on NGM-lite agar medium. Strains were maintained and raised at 20°C, unless otherwise noted. The following mutations were used In this study: LG I: ced-l(e1735), ced-1(n1995) and ced-1(n1506) (Ellis et al, 1991); LG III: dpy-17 (e164) , ced-6(n1813 , n2095), mec-14 (u5 5), ncl-1(e18 6 5) ced7 (n199 7), ced-7(n1892), ced-7(n1996) (Ellis et al, 1991 ), unc-36(e251) (Brenner, 1974) and sDp3(lI1, J) (Rosenbluth et al, 1985); on LG IV: ced-2(e1752) (Hedgecock et al, 1983), ced-5(n1812) and ced-10(n1993) (Ellis et al, 199l). All mutations are described in Hodgkin (1997). EXAMPLE 1 Analysis and Quantifying of Engulfment Animals were anesthetized with 30mM NaN3 and mounted on agar pads to observation using Normarski optics microscope (Sulston & Horvitz, 1977; Avery and Horvitz, 1987). To quantify engulfment of cell corpses generated during embryonic development, the number of persistent cell corpses that were visible in the head region of young L1 larvae that still had only four cells in gonad (i.e., had hatched in the previous four hours) were scored. To quantify the germ line engulfment defect, cell corpses visible within both the distal arm (where the germ cell deaths occur) and the proximal arm (where persistent germ cell corpses can sometimes be observed as they are swept along by the developing oocytes) were counted. EXAMPLE 2 Germline Transformation and Genomic Rescue of ced-6 Transgenic animals were generated using the germline microinjection procedure developed by Mello et al. Cosmids W03A5, F20F 10, F48E8, R02F2, W02G12, T06H6, C48E6, C44D7, F56D2, F43F12, C05D2, T06C9, C05H8 were injected, either singly or in groups (final concentration 20ng/ul for each cosmid), into ced-6(nl813) animals. Plasmid pRF4 was used (final concentration 50-80 ng/ul) as the dominant co-injection marker (Mello et al., 1991); pRF4 carries the mutated collagen gene rol-6(sulOO6gf) and confers a dominant roller (Rol) phenotype. Transgenic lines carrying stably transmitting extrachromosomal arrays were kept for further analysis. To assay for rescue, three-fold embryos laid by transgenic animals were examined for cell corpses under Normaski optics. Transgenic lines that generated embryos with fewer or no corpses were considered to be rescued. To further define the position ofced-6 within F56D2, a number of deletion constructs were created and other fragments subcloned into pBluescript SK(+) II. 50-90 ng/ul of these clones were co-injected with 80-100 ng/ul pRF4 injection marker into ced-6(n1813) worms, and their rescuing ability tested as described above. EXAMPLE 3 Isolation of ced-6 cDNAs To isolate full-length ced-6 cDNAs, a mixed-stage C.elegans lambda Zap cDNA library was screened (gift ofR. Barstead, Oklahoma Medical Research Foundation, Oklahoma City, OK) using established protocols (Sambrook et al., 1989). 3ZP-labeled probe was made using the rescuing 10 kb Xho I genomic fragment as template. Positive phage were transformed into plasmid clones using the in vivo excision protocol. The clones representing F56D2.7 gene from isolated plasmid clones were identified on a Southern blot. For this purpose a 3Zp-labeled probe was generated from RT-PCR product, which represents three exons of predicted F56D2.7. Primers used for RT-PCR: GAATGTTCTCATTTATTG (SEQ ID NO.: 29) and GGATTCAAACGATCCGATG. (SEQ ID NO: 17) From about 300,000 plaques 10 plasmid clones corresponding F56D2.7 cDNAs were isolated. These clones were sequenced for both ends of the insert using the flanking T3 and T7 primers. Two clones with partial SL2 sequence at the 5' end and intact poly(A) tail were identified as full-length F56D2.7 cDNAs. Analysis of these sequence results and the pattern of restriction digestion by Sau3A I also suggested that these clones represent for one transcript. EXAMPLE 4 Reverse transcription-PCR Reverse transcription (RT)-PCR experiments were performed to determine the 5'end of transcripts detected or predicted within the rescuing Xho I genomic fragment. Reverse transcription was performed with following primers: C05D2.6a: GAATCTGTCCATCGCATTGC (SEQ ID NO.: 18), GAATTTCTTTGGGTAGACA (SEQ ID NO.: 19); C05D2.6b: GCTCTGA.AGAACTGTGA (SEQ ID NO.: 20), GACGAGGTGAAGCGATTGTG (SEQ ID NO.: 21); F56D2.7: GGGATCAAACGAATCATC (SEQ ID NO.: 22). These primers were then used in combination with SL1 (GTTTAATTACCCA.AGTTTGAG (SEQ ID NO.: 23)) or SL2 (GGTTTTAACCCAGTTACTCAAG (SEQ ID NO.: 24)) primers for subsequent PCR amplification. Total C. elegans mixed stage RNA was isolated as described previously. RT-PCR was performed using the Superscript Preamplication System (Gibco BRL). EXAMPLE Identification of ced-6 Mutations To determine whether either ced-6 allele resulted in a large physically detectable polymorphism, we generated Southern blots of N2, ced-6(n1813), and ced-6(n2095) genomic DNA digested with various restriction enzymes. A probe generated from the rescuing Xho I genomic fragment detected noval allele-specific bands in ced-6(n2095) using four different restriction enzymes. Analysis of the novel restriction patterns in ced-6(n2095) indicates that this allele carries a complex rearrangement in this region, that covers at least part of F56D2.7, but does not affect the neighboring C05D2.6b transcript. To identify point mutations within F56D2.7, overlapping fragments of the F56D2.7 locus from N2, ced-6(n1813), and ced-6(n2095) mutants were PCR amplified and directly sequenced using the PCR Product Sequencing Kit (Amersham). The overlapping PCR fragments covered the entire F56D2.7 transcription unit and about 1 kb of upstream genomic sequence. Sequences of the primers used for PCR amplification and sequencing are available upon request. EXAMPLE 6 Heat Shock Experiments To test whether ced-6 cDNA can rescue the engulfment defect, Kpn I/Sal I fragment of full-length F56D2.7 cDNA was inserted in Kpn I/Sac I site of MCS II of both pPD49.78 and pPD49.83 vectors which carry hspl6-2 and hspl6-41 promoters, creating the constructs pLQhsl and pLQhs2. The two constructs were co-injected, at 50ng/ul each with 80ng/ul pRF4, to generated stably transmitting extrachromosomal arrays. For our control experiments, we used pPD50.21 and pPD50.15, two derivatives ofpPD49.78 and pPD49.83 in which the lacZ open reading frame has been placed under heat shock promoters. Transgenic lines carrying these constructs were generated as described above. To overexpress ced-6 before cell death occurs during embryonic development, adult animals were put on a plate seeded with E.coli and allowed to lay eggs for one hour. Plates were subsequently parafilmed and subjected to heat shock by transfer to 33°C waterbath for 45 minutes. Following a 75-minute recovery at 20°C, adult animals were removed from the plates. 12-14 hours after heatshock, hatching L1 larvae were scored for corpses in the head region. To overexpress ced-6 after the formation of cell corpses during embryonic development, worm plates containing embryos at all developmental stages (but not larvae) were parafilmed and subjected to heat shock in a 33°C waterbath for minutes. Three hours after the heat shock, freshly hatched L1 larvae were scored for corpses in the head region. To determine the effect of ced-6 overexpression before ce!l death occurs on the engulfment of dying germ cells, L4 stage transgenic animals were transferred to new plates and stored at 20°C. Starting 24 hours after the L4 molt, the worm plates were parafilmed and heat shocked for 45 minutes at 33°C as described above. Animals were examined for germ cell corpses at 12 hours after heat shock, also 18, 24, 36, and 60 hours after heat shock. To overexpress ced-6 after the formation of germ cell corpses, L4 stage transgenic animals were collected and put into several plates, a few for each plate. 24 hours after the L4 molt one plate of worms were heat shocked for 45 minutes as described above. Similarly, 36, 42, 48 and 60 hours after the L4 molt, each plate of worms at one time point were treated with heat. Animals were examined for germ cell corpses 12 hours after heat shock. To overexpress ced-6 in the background of other engulfment mutants, the ced-6 or lacZ-expressing extrachromosomal arrays were transferred from ced-6(nl 813) to a wild-type background, and crossed subsequently to ced-l(e1735) ced-1(n1506), ced-1(n1995), ced-7(1892), ced7(n1996), Ced7(n1997), ced-2(n 1752), ced-5(n 1812) or ced10(n 1993) to generate the corresponding transgenic mutant strains. Heat shock experiments were performed as described above. EXAMPLE 7 Genetic Mosaic Analysis 1000 dpy-17(e164) ced-6(nl 813) mec-14(u5 5) ncl-l (e1865) unc-36(e25) III," sDp3(III, f) were put in worm plates individually. The progenies of these animals were examined to identify animals who laid only DPY UNC progenies under the dissecting microscope. The adult animals were examined under the Normaski Optics immediately after being identified. First the somatic sheath cells were examined, then the body wall muscle descended from D and C lineages. When all body wall muscle cells displayed wild-type, the duplication is lost in P4 lineage. When body wall muscle cells from D lineage are wild-type, while those from C lineage exhibit ncl phenotype, the duplication must be lost from P3 lineage. When body wall muscle cells from both D and C lineages show the ncl phenotype, the duplication must be lost from P2 lineage. The cell corpse in both arms of gonad were also examined for the engulfment phenotype. To find the animals with the duplication lost in the somatic sheath cells, but not in germ cells, dpy-i 7(e164) ced-6(n1813) mec-14(u55) ncl-l (e1865) unc-36(e25) III; sDp3(III, D animals were examined under the Normaski Optics for the loss of the duplication in somatic sheath cells. At the same time cell corpses in gonad were also examined for the engulfment phenotype. EXAMPLE 8 Identification of a human homologue of CED-6 Extensive searches (tblastn) with the ced-6 sequence (Figure 18 Consensus DNA Sequence ofhCED-6) against the public domain databases (EST, Genbank, EMBL, Swissprot and PIR) revealed statistically significant homologies to some ESTS at the carboxyterminal region of the protein (AA443368, AA431995, R33389,R53881). One EST (T48513) showed homology to the Carboxyterminal of the PTB domain and the beginning of the charged region. For 5' RACE analyses a Marathon-ready cDNA colorectal adenocarcinoma, library was used from Clontech. The position of the primers used for RACE and sequencencing is indicated in figure 18. By subsequent cloning and sequence analysis additional sequence information was obtained. Using this additional sequence information and subsequent rounds of database searching (blastn) revealed additional EST, which enabled us to construct a consensus of approx 2400 hp. This sequence was further extended and verified by colony hybridization and sequencing additional RACE products. EXAMPLE 9 RNA Blots (see Figure 25 expression pattern ofhCED-6 in normal human tissues and cancer cell lines by Northern blotting A) Human Multiple Tissue Northern (MTN) Blot B) Human Multiple Tissue Northem (MTN) Blot II C) Human Cancer Cell Line Multiple Tissue Northern (MTN TM) Blot) A Human multiple tissue Northern (MTN-1, Clontech) containing in each lane 2 mg ofpoly A + RNA from eight different human tissues (heart, brain, placenta, lung, liver skeletal muscle, kidney, and pancreas) and a MTN-II human multiple tissue Northern, containing in each lane 2 mg ofpoly A + RNA from spleen, thymus, prostate, testis, ovary, small intestine, colon and peripheral leukocyte, were hybridized according to the manufacturer's instructions and washed out in 0.1 x SSC, 0.2% SDS at 55°C. Also from Clontech, a poly A + RNA blot from human cancer cell lines (melanoma G361, lung carcinoma A549, coloreetal adenocarcinoma SW480, Burkitt's lymphoma Raji Leukemia Molt 4, lympohoblastic leukemia K562, HeLa $3 and promyelocytic leukemia HL60) was tested. EXAMPLE Isolation of the full-length human ced-6 cDNA. Several human EST clones including hbc3123 have been identified through searching variety of database. The hbc3123 EST clone was completely sequenced. One pair of primers, P (ACAATTGCCAGCTTCATAG; SEQ ID NO.: 30) and Q (CTGTTTTCTTGTTTCA.ACATC; SEQ ID NO.: 31) have been designed on the region of PTB domain and subsequently tested for their specificity using human genomic DNA as a template. The result showed that the primers are specific. One set of gtl 0 cDNA libraries (purchased from Clontech) including Brain, Heart, Kidney, Liver, Lung, Pancreas, Placenta, Skeletal Muscle tissues were tested using primers P and Q to detect whether ced-6 is expressed in any of these tissues. The primer Q and a primer against Lgt 10 vector were used to isolate several PCR fragments using brain and pancreas cDNA libraries. These PCR fragments were reamplified using the same primer set and sequenced. The sequence analysis suggested that these PCR fragments allows the extension ofcDNA 130bp upstream of the initiation codon of human ced-6 coding region. The longest PCR fragment was then sent to human EST database to search for more EST clones which have overlap with the isolated PCR fragments but not the hbc3123 EST clone. The Genbank names of these three EST clones are R65982, R65983 and AA159394, respectively. These 3 ESTs together with the PCR fragment and hbc3123 constitute the full-length coding sequence of human CED-6 and about 450 bp of 5'UTR. The human ced-6 cDNA sequenced is confirmed correctly by the sequencing data of hbc3123 EST clone, the sequencing data of the isolated PCR fragments and the sequence data of the many EST clones on the human cDNA region from human EST project. These human ced-6 cDNA data have suggested and guided any experiments shown in both Example 8 and Example 9. See Figure 32. EXAMPLE 11 Human Tissue Distribution of Human CED-6 This is a further example of the human tissue distribution. Two primers against the PTB domain were used to detect whether the cDNA libraries contained human ted-6. The two primers have been tested using human genomic DNA as a template and they axe specific since no background amplification was detected. The result of this tissue distribution study is as follows: I. Information obtained from cDNA library Tissue Presence of human ced-6 cDNA Brain ++ Heart ++ Kidney ++ Liver + Lung ++ Pancreas ++ Placenta ++ Skeletal muscle ++ II. Information obtained from human EST project Tissue EST clones from human EST project Brain 2 Testis 3 Pancreas 4 HCC cell line 1 Aorta 1 Placenta 13 Fetus 1 Pooled sample 2 EXAMPLE 12 The technique known as FISH was carried out, the human ced-6 gene was localized to chomosomal position 2q32.3-q33. EXAMPLE 13 Functional conservation between C. elegans and human ced-6 homologues; overexpression of hCED Rescues the Engulfment Defect of CED-6 Mutants in c. elegans: Given that signal transduction pathways are usually conserved through evolution, it is thought that the human ced-6 homologue (hereatter referred to as hced-6 which encompasses hl CED-6 and/or h3CED-6) might also be involved in promoting the phagocytic removal of apoptotic cells in mammals. To address this question, we tested the human and worm ced-6 genes for functional conservation by overexpressing hced-6 in C. elegans and determining whether it could functionally substitute for the endogenous ced-6 gene. It is shown herein that overexpression of a C. elegans ced-6 cDNA under the control of the heat shock promoters hsp16-2 and hsp16-41 efficiently rescues the engulfment defect in transgenic ced-6 mutant embryos. The same assay was used to test hced-6 for biological activity in C. elegans: constructs were created carrying the hced-6 open reading frame under the control ofhspl6-2 and hsp16-41, and ced6(n1813) mutant animals transgenic for both constructs were tested for rescue of the engulfment defect in late embryos and young larvae. It was found that heat-shocked embryos laid by transgenic mothers, but not non-heat shocked embryos, contained few cell corpses (Figure 31A). These observations suggest that hced-6 can substitute, albeit weakly in the current assay, for C. elegans ced-6, supporting the concept that C. elegans and human ced-6 are functionally conserved. Further assessment as shown in Example 13, showed successful rescue. Partial rescue, or even absence of rescue in certain assays, has been observed previously, even in cases where functional conservation has been established. For example, Wu and Horvitz (1998a) Nature 1998a 3..9.2 501-504, have found that DOCK180, the mammalian homologue ofC. elegans CED-5, efficiently rescued the distal tip cell migration defect of CED-5 mutants, but not the engulfment defect. Experimental Procedures The open reading flame of hced-6 was PCR-amplified using oligonucleotides flanking the start and stop codons, and subcloned into the heat shock vectors pPD49.78 and pPD49.83, previously digested with Kpn I and Sac I (see before). The two constructs were then injected into ced-60H 813) animals as described previously to establish stably transmitting transgenic lines. To score for rescue of the engulfment defect in embryos and in the adult germ line, transgenic animals were submitted to heat-shock and the number of cell corpses quantified as described previously herein. Table 2 Overexpression of human ced-6 homologue reduces the number of persistent cell corpses in ced-6(n1813) late embryos. Genotype Wild Type (N2) ced-6(n1813)++-tced-6(n1813); hs: :hced-6 Ill Persistent cell corpses heatshock + heatshock -Jr" One of the isolated PCR fragments was fused to the hbc3123 EST clone. pLQhced-6.1, the fusion cDNA, has 130 nucleotides upstream of the initiation codon ATG. Two primers, Hhsl (GGGGTACCGAATTCTGATGGCA.AC; (SEQ ID NO.:27)) and Hhs3 (CGAGCTCGATCAATAGTGAAGGTGAGG; (SEQ ID NO.: 28)) were used to amplify the open reading frame of human ced-6 eDNA. The PCR fragment was digested subsequently with Kpn I and Sac I, and inserted into Kpn I and Sac I sites of both ppD49.78 and ppD49.83 heat shock vectors. The heat shock constructs, pLQhsl and pLQhs2, 50 ng/p.l for each, whre then co-injected with a marker pRF4 (80 ng/ tl) into the germline of adult ced-6(h1813) hermaphrodites. nced-6 was examined for its ability to rescue the engulfment defect in embryo progeny of ced-6(nl813) transgenic animals following an established procedure, as described herein. The rescuing ability of hCED-6 for the engulfment defect of ced-6(n1813) in the adult gonas was also tested. Transgenic animals at L4/adult molt were picked and put on a fresh plate. 36 hours later these animals were treated with a 45 minute heat shock at 33 °C. Twelve hours after the heat shock, cell corpses were scored in one gonad arm. Control experiments, such as transgenic animals withut heat treatment, ced-6(n1813) animals at the same development stage with or without heat shock, were also used. These experiments show that overexpression of hced-6 rescued the engulfment defect of CED-6 mutants in C. elegans in a germ line. These experiments confirm that human ced-6's (e.g., h3CED-6) function induces the phagocytosis of apoptotic cells. Figures 31A and 31B. EXAMPLE 14 Sequences can be obtained in both deposits using T3 or T7 primers (either one or both can be used, they are at different sites of the actual insert). Both are commercially available from Clontech (#1227 and #1228) and sequence is shown below T7 primer: 5'(TAATACGACTCACTATAGGGAGA)3'(SEQ ID NO.: 25) T3 primer: 5'(ATTAACCCTCACTAAAGGGA)Y(SEQ ID NO.: 26) REFERENCES: Avery, L., and Horvitz, H. (1987). A cell that dies during wild-type C. elegans development can function as a neuron in a ced-3 mutant. Cell 51, 1071-1078. Blaikie, P., Immanuel, D., Wu, J., Li, N., Yajnik, V., and Margolis, b. (1994). A region in Shc distinct from the SH2 domain can bind tyrosine-phosphorylated growth factor receptor. The Journal of Biological Chemistry 269, 32031-32034. Blumenthal, T. (1995). Trans-splicing and polycistronic transcription in Caenorbabditis elegans. TIG 11,132-136. Blumenthal, T., and Steward, K. (1997). RNA processing and gene structure. In C. ELEGANS II, D. L. Riddle, T. Blumenthal, B. J. Meyer and J. R. Priess, eds. (Cold Spring Harbor: Cold Spring Harbor Laboratory Press), pp. 117-145. Bork, P., and Margolis, B. (1995). A phosphotyrosine interaction domain. Cell 80, Brennar, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71-94. Driscoll, M., and Chalfie, M. (1991). The mec-4 gene is a member of a family of Caenorhabditis elegans genes that can mutate to induce neuronal degeneration. Nature 349, 588-593. Driscoll, M. (1992). Molecular genetics of cell death in the nematode Caenorhabditis elegans. The Journal of Neurobiology 23, 1327-1351. Duvall, E., Wyllie, A. H., and Morris, R. G. (1985). Macrophage recognition of cells undergoing programmed cell death (apoptosis). Immunology 56, 351-358. Ellis, R. E., Jacobson, D. M., and Horvitz, H. R. (1991). Genes required for the engulfment of cell corpses during programmed cell death in Caenorhabditis elegans. Genetics 129, 79-94. Fadok, V. A., Savill, J. S., Haslett, C., Bratton, D. L., Doherty, D. E., Campbell, P. A., and Henson, P. M. (1992). Different populations of macrophages use either the vitronectin receptor or the phosphatidylserine receptor to recognize and remove apoptotic cells. The Journal of Immunology 149, 4029-4035. Fadok, V. A., Voelker, D. R., Campbell, P. A., Cohen, J. J., Bratton, D. L., and Henson, P. M. (1992). Exposure of phosphatidylserine on the Surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. The Journal of Immunology 148, 2207-2216. Finan, P., Shimizu, Y., Gout, I., Hsuan, J., Truong, O., Butcher, C., Bennett, P., Waterfield, M. D., and Kellie, S. (1994). An SH3 domain and proline-rich sequence mediate an interaction between two components of the phagocyte NADPH oxidase complex. The Journal of Biochemical Chemistry 269, 13752-13755. Franc, N. C., Dimarcq, J.-L., Lagueux, M., Hoffmann, J., and Ezekowitz, R. A. B. (1996). Croquemort, A novel Drosophila hemocyte/macrophage receptor that recognizes apoptotic cells. Immunity 4, 431-443. Frise, E., Knoblich, J., Younger-Shepherd, S., Jan, L., and Jan, Y. (1996). The Drosophila Numb protein inhibits signaling of the Notch receptor during cell-cell interaction in sensory organ lineage. Proc Natl Acad Sci U S A 93(15), 11925-11932. Geer, P. v. d., and Pawson, T. (1995). The PTB domain: a new protein module implicated in signal transduction. TIBS 20, 277-280. Gout, I., Dhand, R., Hiles, I. D., Truong, O., Totty, N. F., Hsuan, J., Booker, G. W., Campbell, L. D., and Waterfield, M. D. (1993). The GTPase dynamin binds to and activated by a subset of SH3 domains. Cell 75, 25-36. Grabs, D., Slepnev, V. I., Songyang, Z., David, C., Lynch, M., Cantley, L. C., and Camilli, P. D. (1997). The SH3 domain of amphiphysin binds the proline-rich domain of dynamin at a single site that defines a new SH3 binding consensus sequence. The Journal of Biological Chemistry 272, 13419-13425. Greenberg, S., Chang, P., and Silverstein, S. (1993). Tyrosine phosphorylation is required for Fc receptor-mediated phagocytosis in mouse macrophages. J Med Med Greenberg, S., Chang, P., and Silverstein, S. (1994). Tyrosine phosphorylation of the gamma subunit of Fc gamma receptors, p72syk, and paxillin during Fc receptor-mediated phagocytosis in macrophages. J Biol Chem 269, 3897-3902. Greenberg, S. (1995). Signal transduction of phagocytosis. Trends in Cell Biology 5, 93-99. Grigg, J., Savill, J., Sarraf, C., Haslett, C., and Silverman, M. (1991). Neutrophil apoptosis and clearance from neonatal lungs. Lancet 338, 720-722. Hedgecock, E. M., Sulston, J. E., and Thomson, J. N. (1983). Mutations affecting programmed cell deaths in the nematode Caenorhabditis elegans. Science 220, 1277-1279. Hedgecock, E. M., and Herman, R. K. (1995). The nuc-1 gene and genetic mosiac of Caenorhabditis elegans. Genetics 141,989-1006. Hengartner, M. O., and Horvitz, H. R. (1994). C. elegans cell survival gene ced-9 encodes a functional homologue of the mammalian proto-oncogene bcl-2. Cell 76, 665-676. Hengartner, M. O. (1997). Cell Death. In C. ELEGANS II, D. L. Riddle, T. Blumenthal, B. J. Meyer and J. R. Priess, eds. (Plainview: Cold Spring Harbor Laboratory Press), pp. 383-415. Herman, R. K. (1984). Analysis of genetic mosiac of the nematode Caenorhabditis Elegans. Genetics 108, 165-180. Hirsh, D., Oppenheim, D., and Klass, M. (1976). Development of the reproductive system of Caenorhabditis elegans. Developmental Biology 49, 200-219. Horvitz, J. E. S. a. H. R. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Developmental Biology 56, 110-156. Huang, X.-Y., and Hirsh, D. (1989). A second trans-spliced RNA leader sequence in the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 86, 8640-8644. Kavanaugh, W. M., and Williams, L. T. (1994). An alternative to SH2 domains for binding tyrosine-phosphorylated proteins. Science 266. Kavanaugh, W. M., Turck, C. W., and Williams, L. T. (1995). PTB domain binding to signaling proteins through a sequence motif containing phosphotyrosine. Science 268, 1177-1179. Kimble, J., and Hirsh, D. (1979). The postembryonic cell lineages of the hermaphrodite and male gonads in Caenorhabditis elegans. Developmental Biology 70, 396-417. Koch, C. A., Anderson, D., Moran, M. F., Ellis, C., and Pawson, T. (1991). SH2 and SH3 Domains: elements that control interactions of cytoplasmic signaling proteins. Science 252, 252-673. Kramer, J. M., French, R. P., Park, E., and Johnson, J. J. (1990). The Caenorhabditis elegans rol-6 gene, which interacts with the sqt-1 collagen gene to determine organismal morphology, encodes a collagen. Molecular and cellular Biology 10, Lockshin, R. A. (1981). Cell death in metamorphosis. In Cell death in biology and pathology, R. A. Lockshin and I. D. Browen, eds. (London: Chapman & Hall), pp. 79-122. Luciani, M.-F., and Chimili, G. (1996). The ATP binding cassette transporter ABC1, is required for the engulfment of corpses generated by apoptotic cell death. The EMBO Journal 15,226-235. Mello, C., and Fire, A. (1995). DNA transformation. In Methods in Cell Biology, H. F. Epstein and D. C. Shakes, eds. (San Diego: Academic Press), pp. 452-482. Morris, R. G., Duvall, E., Hangreaves, A. D., and Wyllie, A. H. (1984). Hormone-induced cell death. 2. Surface changes in thymocytes undergoing apoptosis. American Journal of Pathology 115,426-436. Morris, R. G., Hargreaves, A. D., Duva!l, E., and Wyllie, A. H. (1984). Hormone-Induced Cell Death.. Pawson, T., and Schlessinger, J. (1993). SH2 and SH3 domains. Current Biology 3, 434-442. Platt, N., Suzuki, H., Kurihara, Y., Kodama, T., and Gordon, S. (1996). Role for the class A macrophage scavenger receptor in the phagocytosis of apoptotic thymocytes in vitro. Proc. Natl. Acad. Sci. USA 93, 12456-12460. Ravetch, J. V. (1994). Fc Receptor: Rubor Redux. Cell 78, 553-560. Ren, Y., Silverstein, R. L., Allen, J., and Savill, J. (1995). CD36 Gene Transfer Confers Capacity for Phagocytosis of Cells Undergoing Apoptosis. J. Exp. Med. 18, 1857-1862. Roberson, A., and Thomson, N. (1982). Morphology of programmed cell death in the ventral nerve cord ofCaenorhabditis elegans Larvae. J. Embryol.Exp.Morph 67, 89-100. Rosenbluth, R. E., Cuddeford, C., and Baillie, D. L. (1985). Mutagenesis in Caenorhabditis elegans. II. A spectrum of mutational events induced with 1500 R of r-radiation. Genetics 109, 493-511. Rosenshine, I., and Finlay, B. B. (1993). Exploitation of Host Signal Transduction Pathways and Cytoskeletal Functions by Invasive Bacteria. BioEssays 15, 17-24. Rotello, R. J., Fernandez, P.-A., and Yuan, J. (1994). Anti-apogens and anti-engulfens: monoclonal antibodies reveal specific antigens on apoptotic and engulfment cells during chicken embryonic development. Development 120, 1421-1431. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular cloning: a laboratory manual (Cold Spring Harbor, NY: Cold Spring Harbor Press).. Savill, J., Wyllie, A., Henson, J., Walport, M., Henson, P., and Haslett, C. (1989). Macrophage phagocytosis of aging neutrophils in inflamation. Programmed cell death in the neutrophil leads to its recognition by macrophages. J Clin Invest 83, Savill, J., Dransfield, I., Hogg, N., and Haslett, C. (1990). Vitronectin receptor-mediated phagocytosis of cells undergoing apoptosis. Nature 343, 170-173. Savill, J., Hogg, N., Ren, Y., and Haslett, C. (1992). Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J Clin Invest 90, 1513-1522. Savill, J., Fadok, V., Henson, P., and Haslett, C. (1993). Phagocyte recognition of cells undergoing apoptosis.. Songyang, Z., and Cantley, L. C. (1995). Recognition and specificity in protein tyrosine kinase-mediated signaling. TIBS 20, 470-475. Songyang, Z., Margolis, B., Chaudhuri, M., Shoelson, S. E., and Cantley, L. C. (1995). The phosphotyrosine interaction domain of SHC recognizes tyrosine-phosphorylated NPX'Y motif. The Journal of Biological Chemistry 270, 14863-14866. Spieth, J., Brooke, G., Kuersten, S., Lea, K., and Blumenthal, T. (1993). Operons in C. elegans: polycistronic mRNA precursors are processed by trans-splicing of SL2 to downstream coding region. Cell 73, 521-532. Stinchcomb, D. T., Shaw, J. E., Cart, S. H., and Hirsh, D. (1985). Extrachromosomal DNA transformation of Caenorhabditis elegans. Molecular and Cellular biology 5, 3484-3496. Stringham, E. G., Dixon, D. K., Jones, D., and Candido, E. P. M. (1992). Temporal and spatial expression patterns of the small heat shock (hsp 16) genes in transgenic Caenorhabditis elegans. Molecular Biology of the Cell 3, 221-233. Sulston, J. (1976). Post-embryonic development in the ventral cord of Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci 275, 287-297. Sulston, J. E., and Horvitz, H. R. (1977). Post-embryonic Cell Lineages of the Nematode, Caenorhabditis elegans. Developmental Biology 56, 110-156. Sulston, J. E., Schierenberg, E., White, J. G., and Thomson, J. N. (1983). The embryonic cell lineage of the nematode Caenorhabditis elegans. Developmental Biology 100, 64-119. Wilson, R., Ainscough, R., Anderson, K., Baynes, C., Berks, M., Bonfield, J., Burton, J., Connell, M., Copsey, T., Cooper, J., Coulson, A., Craxton, M., Dear, S., Du, Z., Durbin, R., Favello, A., Fraser, A., Fulton, L., Gardner, A., Green, P., Hawkins, T., Hiller, L., Jier, M., Johnston, L., Jones, M., Kershaw, J., Kirsten, J., Laisster, N., Latrellie, P., Lightning, J., Lloyd, C., Mortimore, B., O'Callaghan, M., Parsons, J., Percy, C., Rifken, L., Roopra, A., Saunders, D., Shownkeen, R., Sims, M., Smaldon, N., Smith, A., Smith, M., Sonnhammer, E., Staden, R., Sulston, J., Thierry-Mieg, J., Thomas, K., Vaudin, M., Vaughan, K., Waterston, R., Watson, A., Welnstock, L., Wilkinson-Sproat, J., and Wohldman, P. (1994). 2.2 Mb of contiguous nucleotide sequence from chromosome [II of C. elegans. Nature 368, 32-38. Wolf, G., Trub, T., Ottinger, E., Groninga, L., Lynch, A., White, M. F., Miyazaki, M., Lee, J., and Shoelson, S. E. (1995). PTB domain of IRS-1 and Shc have distinct but overlapping binding specificities. The Joumal of Biological Chemistry 270, 27407-27410. Wyllie, A. H., Kerr, J. F. R., and Currie, A. R. (1980). Cell Death: The significance of apoptosis. International Review of Cytology 68, 251-306. Xu, X.-X., Yang, W., Jackowski, S., and Rock, C. O. (1995). Cloning of a novel phosphoprotein regulated by colony-stimulating factor 1 shares a domain with the Drosophila disabled gene product. The Journal of Biological Chemistry 270, 14184-14191. Yenush, L., and White, M. F. (1997). The IRS-signalling system during insulin and cytokine action. BioEssay 19, 491-500. Yu, H., Chen, J. K., Feng, S., Dalgamo, D. C., Brauer, A. W., and Schreiber, S. L. (1994). Structural basis for the binding ofproline-rich peptides to SH3 domains. Cell 76, 933-945. Zhou, M.-M., Ravichandran, K. S., Olejniczak, E. T., Petros, A. M., Meadows, R. P., Sat-tler, M., Harlan, J. E., Wade, W. S., and Fesik, S. J. B. S. W. (1995). Structure and ligand recognition of the phosphotyrosine binding domain of Shc. Nature 378, 584-592. Zorio, D. A. R., Cheng, N. N., Blumenthal, T., and Spieth, J. (1994). Operons as a common form of chromosomal organization in C. elegans. Nature 372, 270-272. While this invention has been particularly shown and described with references to preferred embodiments thereof, it will be understood by those skilled the art that various changes in form and details may be made therein without departing from the spirit and scope of the invention as defined by the appended Throughout this specification and the claims which follow, unless the context requires otherwise, the word "comprise", and variations such as "comprises" and "comprising", will be understood to imply the inclusion of a stated integer or step or group of integers or steps but not the exclusion of any other integer or step or group of integers or steps. The reference to any prior art in this specification is not, and should not be taken as, an acknowledgment or any form of suggestion that that prior art forms part of the common general knowledge in Australia. (57) Abstract The invention relates to a signal transduction pathway which promotes phagocytosis of 0 10 20 30 i , apoptotie cells and in particular relates to a protein known as CED--6 in the nematode worm C. elegans, human equivalents of said protein and nucleic acids encoding them. The invention too also relates to use of the proteins and encoding nucleic acids in assay methods for detecting compounds which enhance or inhibit the aforesaid signal transduction pathway and use of the 2OO proteins, nucleic acids and identified enhancer or inhibitor compounds in methods of treatment of human or animal disease. CLAIMS DEFINING THE INVENTION ARE AS FOLLOWS:
An isolated protein selected from the group consisting of:
a) a protein comprising the amino acid sequence as shown in SEQ ID No. 2, b) a protein comprising the amino acid sequence as shown in SEQ ID No. 4, c) a protein comprising the amino acid sequence as shown in SEQ ID No. 6, d) a protein comprising the amino acid sequence as shown in SEQ ID No. 8, e) a protein comprlsmg the amino acid sequence as shown in SEQ ID No. 10, f) a proteiia comprising the ammo acid sequence as shown in SEQ ID No. 12, g) a protein comprising the amino acid sequence as shown in SEQ ID No. 14, h) a protein comprising the amino acid sequence as shown in SEQ ID No. 16, i) a protein comprising an ammo acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No, 2, 4, 6, 8, 10, 12, 14, or 16, and J) a protein comprising the amino acid sequence encoded by a nucleic acid molecule comprising the nucleic acid sequence as shown in SEQ ID Nos. 1, 3,5,7,9, 11, 13or An isolated nucleic acid molecule selected from the group consisting of:
a) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 1, b) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 3, c) a nucleic acid molecule comprising the nucleic acid sequence shown in d) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 9, e) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 11 f) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 13, g) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 15, h) a nucleic acid molecule comprising the complement of the nucleic acid sequence shown in SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15, i) a nucleic acid molecule capable of hybridizing to a nucleic acid having a nucleic acid sequence according to (h) under conditions of high stringency, j) a nucleic acid molecule which encodes an amino acid which is at least 70% identical to an amino acid having the amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16, and k) a nucleic acid molecule having a nucleic acid sequence which is at least 70% identical to the nucleic acid sequence of SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15.
An expression vector comprising a nucleic acid molecule selected from the group consisting of:
a) a nucleic acid molecule comprising the nucleic acid sequence shown in b) a nucleic acid molecule comprising the complement of the nucleic acid sequence shown in SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15, c) a nucleic acid molecule capable of hybridizing to a nucleic acid having a nucleic acid sequence according to (b) under conditions of high stringency, d) a nucleic acid molecule which encodes an amino acid which is at least 70% identical to an amino acid having the amino acid sequence of SEQ ID Nos.
2, 4, 6, 8, 10, 12, 14, or 16, and e) a nucleic acid molecule which encodes an amino acid having an amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16.
The expression vector of claim 3, comprising DNA encoding a reporter gene positioned in said vector such that expression of said nucleic acid results in expression of said reporter gene.
The expression vector of claim 4, wherein said reporter gene encodes green fluorescent protein.
A mammalian cell-line transfected with a nucleic acid selected from the group consisting of:
a) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No. 1, 3, 5, 7, 9, 11, 13, or 15, b) a nucleic acid molecule comprising the complement of the nucleic acid sequence shown in SEQ ID No. 1, 3, 5, 7, 9, 11, 13, or 15, c) a nucleic acid molecule capable of hybridizing to a nucleic acid having a nucleic acid sequence according to (b) under conditions of high stringency, d) a nucleic acid molecule which encodes an amino acid which is at least 70% identical to an amino acid having the amino acid sequence of SEQ ID Nos.
2, 4, 6, 8, 10, 12, 14, or 16, and e) a nucleic acid molecule which encodes an amino acid having an amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16.
A mammalian cell-line as claimed in claim 6, which is transfected with an expression vector comprising said nucleic acid and a reporter gene, said reporter gene being positioned in said vector such that expression of said nucleic acid results in expression of said reporter gene.
A mammalian cell-line as claimed in claim 7, wherein said reporter gene encodes green fluorescent protein.
A mammalian cell-line as claimed in claim 6, wherein said cell-line is selected from the group consisting of: a fibroblast cell-line and an epithelial cell-line.
A mammalian cell-line as claimed in claim 6, wherein said cell-line is selected from the group consisting of: COS1, BHK21, L929, CV1, SWISS 3T3, HT144, A mammalian cell-line as claimed in claim 10, which is an MCF7 cell-line.
A mammalian cell-line as claimed in claim 7, which is an MCF7 cell-line.
A mammalian cell-line as claimed in claim 6, wherein said cell-line is a primary cell-line.
A mammalian cell-line as claimed in claim 13 wherein said cell-line is selected from the group consisting of: human dermal FIBs, dermal keratinocytes, leukocytes, monocytes, and macrophages.
A non-human transgenic animal comprising a gene encoding a protein, wherein the protein is selected from the group consisting of:
a) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 2, b) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 4, c) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 6, d) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 8, e) a protein comprismg an amino acid having an amino acid sequence as shown in SEQ ID No. 10, f) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 12, g) a protein comprismg an amino acid having an amino acid sequence as shown in SEQ ID No. 14, h) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 16, i) a protein comprising an amino acid having an amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No. 2, 4, 6, 8, 10, 12, 14, or 16, and J) a protein comprising an amino acid having an amino acid sequence encoded by the nucleic acid molecule having the nucleic acid sequence as shown in SEQ ID No's 1, 3, 5, 7, 9, 11, 13 or A ,transgenic nematode worm which is lacking all or substantially all of the function of its native CED-6 gene and has been transfected or transformed with a nucleic acid selected from the group consisting of:
a) . a nucleic acid mo!e ule comprising the nucleic acid sequence shown in SEQ ID No. 7, 9, 11, 13, or 15, b) a nucleic acid molecule comprising the complement of the nucleic acid sequence shown in SEQ ID Nos. 7, 9, 11, 13, or c) a nucleic acid molecule capable of hybridizing to a nucleic acid having a nucleic acid sequence according to (b) under conditions of high stringency, d) a nucleic acid molecule encoding amino acid which is at least 70% identical to an amino acid having the amino acid sequence of SEQ ID Nos. 8, 10, 12, 14, or 16, and e) a nucleic acid molecule which encodes an amino acid having an amino acid sequence of SEQ ID Nos. 8, 10, 12, 14, or 16.
The transgenic nematode worm of claim 16 which is C. elegans.
A method for determining whether a compound is an inhibitor or an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells which method comprises exposing a transgenic mammalian cell transfected with a nucleic acid selected from the group consisting of:
a) comprising: a protein comprising an amino acid having the amino acid of sequence SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; a nucleic acid molecule that encodes an amino acid having the amino acid sequence of SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; an amino acid having the amino acid sequence encoded by a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15; or a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15, b) a nucleic acid molecule which encodes the amino acid having an amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16, to apoptotic particles and measuring the rate of phagocytic uptake of said particles by said transgenic cells in the presence and absence of said compound, wherein an increased rate of phagocytosis indicates an enhancer and a decreased rate of phagocytosis indicates an inhibitor.
A method as claimed in claim 18, wherein said apoptotic particles are selected from the group consisting of: opsonized apoptotic neutrophils, opsonized apoptotic lymphocytes, opsonized apoptotic erythrocytes, opsonized killed bacteria and opsonized killed yeast.
A method as claimed in claim 19, wherein said apoptotic particles are labelled.
A method as claimed in claim 20, wherein said label is selected from the group consisting of: a non-fluorescent dye, a fluorescent dye, a non-fluorescent dye linked to an antibody and a fluorescent dye linked to an antibody.
A method as claimed in claim 18, wherein the transgenic mammalian cell is a fibroblast cell or an epithelial cell.
A method as claimed in claim 22, wherein the transgenic mammalian cell is selected from the group consisting of: COS1, BHK21, L929, CV1, SWISS 3T3, HT144, IMR32, HEPG2, MDCK, MCF7, 293, Hela, A549, SW48 and G361.
A method as claimed in claim 18, wherein said transgenic mammalian cell is a primary cell.
A method as claimed in claim 24, wherein transgenic mammalian cell is selected from the group consisting of: human dermal FIBs, dermal keratinocytes, leukocytes, monocytes and macrophages.
A method as claimed in claim 21, wherein the phagocytosed apoptotic particles are detected by a method selected from the group consisting of: light microscopy, fluorescence microscopy, quantitative spectrofluorometry, and flow cytometry.
A compound identified by the method of claim 18, as an inhibitor or an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells, when used for treating inflammation, autoimmune disease, cancer, neurodegenerativ e disease, stroke or sickle-cell anemia.
A method for determining whether a compound is an inhibitor or an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells, comprising:
(1) introducing into a mammalian cell a protein selected from the group consisting of:
a) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 2, 4, 6, 8, 10, 12, 14, or 16, b) a protein comprising an amino acid having an amino acid sequence which is at least 70% identical with an amino acid having an amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16, c) a protein comprising the amino acid having an amino acid sequence encoded by a nucleic acid molecule having acid sequence shown in introducing into a mammalian cell a vector expressing RNA antisense that inhibits transcription of a protein in one of the groups (a) to (c) above, and (2) exposing the mammalian cell to apoptotic particles and measuring the rate of phagocytic uptake of said particles by said cell in the presence or absence of said compound wherein an increased rate of phagocytosis indicates an enhancer and a decreased rate of phagocytosis indicates an inhibitor.
A method as claimed in claim 28, wherein said apoptotic particles are selected from the group consisting of: opsonized apoptotic neutrophils, opsonized apoptotic lymphocytes, opsonized apoptotic erythrocytes, opsonized killed bacteria, and opsonized killed yeast.
A method as claimed in claim 28, wherein said apoptotic particles are labelled.
A method as claimed in claim 30, wherein said label is selected from the group consisting of: a non-fluorescent dye, a fluorescent dye, a non-fluorescent dye linked to an antibody, and a fluorescent dye linked to an antibody.
A method as claimed in claim 28, wherein the mammalian cell is a fibroblast cell or an epithelial cell.
A method as claimed in claim 32, wherein the mammalian cell is selected from the group consisting of: COS1, BHK21, L929, CV1, SWISS 3T3, HT144, IMR32, HEPG2, MDCK, MCF7, 293, Hela, A549, SW48 and G361.
A method as claimed in claim 28, wherein said mammalian cell is a primary cell.
A method as claimed in claim 34, wherein said mammalian cell is selected from the group consisting of: human dermal FIBs, dermal keratinocytes, leukocytes, monocytes, and macrophages.
A method as claimed in claim 30, wherein in phagocytosed apoptotic particles are detected by a method selected from the group consisting of: light microscopy, fuorescence microscopy, quantitative spectrofluorometry and flow cytometry.
A compound identified by the method of claim 28 as an inhibitor or an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells, when used for treating inflammation, autoimmune disease, cancer, neurodegenerative disease, stroke or sickle-cell anemia.
A method for determining whether a compound is an inhibitor or an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells which method comprises exposing a mammalian cell selected from the group consisting a transgenic mammalian cell transfected with a nucleic acid selected from the group comprising:
a) a nucleic acid molecule having the nucleic acid sequence shown in SEQ ID No. 1, 3, 5, 7, 9, 11, 13, or 15, b) a nucleic acid complement of a nucleic acid molecule having the nucleic acid sequence shown in SEQ ID No. 1, 3, 5, 7, 9, 11, 13, or 15, c) a nucleic acid molecule capable of hybridizing to a nucleic acid having a nucleic acid sequence according to (b) under conditions of high stringency, a mammalian cell which expresses a protein selected from the group consisting of:
a) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 2, 4, 6, 8, 10, 12, 14, or 16, b) a protein comprising amino acid sequence which is at least 70% identical with SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16; and c) a protein comprising an amino acid having an amino acid sequence encoded by a nucleic acid having a nucleic acid sequence shown in SEQ ID. Nos. 1, 3, 5, 7, 9, 11, 13, or 15, and a mammalian cell which comprises a vector expressing RNA antisense to the DNA encoding a protein selected from groups consisting of:
a) a protein comprising an amino acid having amino acid sequence as shown in SEQ ID No. 2, 4, 6, 8, 10, 12, or 16, b) a protein comprising an amino acid having amino acid sequence which is at least 70% identical with SEQ ID Nos. 2, 4, 6, 8, 10, 12, c) a protein comprising an amino acid having amino acid sequence encoded by a nucleic acid having a nucleic acid sequence shown in SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15, and to a compound to be tested and determining whether there is any change in the organization of the actin cytoskeleton, wherein an increase in the rearrangement of actin cytoskeleton indicates the enhancer, and a decrease in the rearrangement of actin cytoskeleton indicates the inhibitor.
A method as claimed in claim 38, wherein the actin cytoskeleton is visualized with a fluorescent dye which is linked to a compound which interacts with Factin.
A method as claimed in claim 39, wherein said linker compound is phalloyidine.
A method as claimed in claim 38, wherein the transgenic mammalian cell is a fibroblast cell or an epithelial cell.
A method as claimed in claim 41, wherein the transgenic mammalian cell is selected from the group consisting of: COS1, BHK21, L929, CV1, SWISS 3T3, HT144, IMR32, HEPG2, MDCK, MCF7, U293, Hela, A549, SW48, and G361.
A method as claimed in claim 38, wherein said transgenic mammalian cell is a primary cell.
A method as claimed in claim 43, wherein said transgenic mammalian cell is selected from the group consisting of: human dermal FIBs, dermal keratinocytes, leukocytes, monocytes, and macrophages.
A compound identified by the method of claim 38, as an inhibitor or an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells, when used for treating inflammation, autoimmune disease, cancer, neurodegenerative disease, stroke or sickle-cell anemia.
A method for determining whether a compound is an inhibitor or an enhancer of expression of a gene encoding a protein which participates in a signal transduction path way which promotes phagocytosis of apoptotic cells, comprising:
(a) exposing a transgenic mammalian cell as claimed in claim 7 to said compound; (b) measuring the level of expression of said reporter gene; and (c) comparing said expression with the level of expression of said reporter gene in the absence of said compound, wherein an increased rate of phagocytosis indicates an enhancer and a decreased rate of phagocytosis indicates an inhibitor.
A compound identified by the method of claim 46, as an inhibitor or an enhancer of expression of a gene encoding a protein which participates in a signal transduction pathway which promotes phagocytosis of apoptotic cells, when used for treating inflammation, autoimmune disease, cancer, neurodegenerative disease, stroke or sickle-cell anemia.
An antibody directed against an epitope of the protein selected from the group consisting of:
a) a protein compnsang an amino acid having an amino acid sequence as shown in SEQ ID No. 2, b) a protein comprising an amino acid having an amano acid sequence as shown in SEQ ID No. 4, c) a protein comprising an amano acid having an amino acid sequence as shown in SEQ ID No. 6, d) a protean comprising an amino acid having an amano acid sequence as shown in SEQ ID No. 8, e) a protean comprlsang an amino acid having an amino acid sequence as shown in SEQ ID No. 10, f) a protein comprlsang an amino acid having an amino acid sequence as shown in SEQ ID No. 12, g) a protein compnsang an amano acid having an amano acid sequence as shown in SEQ ID No. 14, h) a protein comprising an amino acid having an amino acid sequence as shown in SEQ ID No. 16, i) a protein comprising an amino acid having an amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No. 2, 4, 6, 8, 10, 12, 14, or 16 and J) a protein comprising an amino acid having an amino acid sequence encoded by the nucleic acid molecule having the nucleic acid sequence as shown in SEQ ID No. 1, 3, 5, 7, 9, 11, 13, or An antibody as claimed in claim 48, which is a monoclonal antibody.
A method of treating in an individual having a disease selected from the group consisting of: inflammation, autoimmune disease and cancer comprising administering to a patient a medicament comprising an effective amount of a protein selected from the group consisting of:
a) a protein comprising an amino acid having the amino acid sequence of SEQ ID No: 8, b) a protein comprising an amino acid having the amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No. 8, and c) a protein comprising an amino acid having the amino acid sequence encoded by the nucleotide sequence of SEQ ID No. 7.
A method of treating an individual having a disease selected from the group consisting of: inflammation, autoimmune disease and cancer, comprising administering to a patient in need thereof an effective amount of a compound which is an enhancer of the protein of Claim 1 and is involved in a signal transduction pathway which promotes phagocytosis of apoptotic cells.
A method of treating an individual having a disease selected from the group consisting of: inflammation, autoimmune disease and cancer, comprising administering to a patient in need thereof an effective amount of a compound which is an enhancer of expression of the nucleic acid molecule of Claim 2 and encodes a protein which participates in a signal transduction pathway which promotes phagocytosis of apoptotic cells.
A method of treating an individual having a disease selected from the group consisting of: inflammation, autoimmune disease and cancer, comprising administering to a patient in need thereof an effective amount of a nucleic acid molecule having the nucleic acid sequence selected from the group consisting of:
SEQ ID No. 7, 13, and A method of treating an individual having a disease selected from the group consisting of: neurodegenerative disease, stroke and sickle-cell anemia, comprising administering to a patient in need thereof an effective amount of a protein selected from the group consisting of:
a) a protein comprising an amino acid having the amino acid sequence of SEQ ID No.: 8, 14, 16, b) a protein comprising an amino acid having the amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No.: 8, 14, c) a protein comprising an amino acid having amino acid sequence encoded by a nucleic acid molecule having the nucleic acid sequence of SEQ ID No.: 7, 13, or A method of treating an individual having a disease selected from the group consisting of: neurodegenerative disease, stroke and sickle cell anemia, comprising administering to a patient in need thereof an effective amount of a compound identified as an inhibitor of the protein of Claim 1 and is involved in a signal transduction pathway which promotes phagocytosis of apoptotic cells.
method of treating an individual having a disease selected from a group consisting of: neurodegenerative disease, stroke and sickle cell anemia, comprising administering to a patient in need thereof an effective amount of a compound identified as an inhibitor of expression of the nucleic acid molecule of Claim 2 and encodes a protein which participates in a signal transduction pathway which promotes phagocytosis of apoptotic cells.
A method of treating an individual having a disease selected from the group consisting of: neurodegenerative disease, stroke, and sickle-cell anemia, comprising administering to a patient in need thereof an effective amount of a nucleic acid selected from the group consisting of:
a) a nucleic acid molecule comprising the nucleic acid sequence of SEQ ID b) a nucleic acid molecule which hybridizes to a nucleic acid having a nucleic acid sequence of SEQ ID No. 7, 13, or 15, and c) a nucleic acid molecule that encodes an amino acid having the amino acid sequence of SEQ ID NO.: 8, 14, or 16.
A pharmaceutical composition comprising a protein selected from the group consisting of:
a) a protein comprising an amino acid having an amino acid sequence of SEQ ID No.: 8, 14, or 16, b) a protein comprising an amino acid having an amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No. 8, 14, or 16, and c) a protein comprising an amino acid having amino acid sequence encoded by a nucleic acid having the nucleic acid sequence of SEQ ID No. 7, 13, or 15, and a pharmaceutically acceptable carrier.
A pharmaceutical composition comprising a compound as claimed in claim 27 and a pharmaceutically acceptable carrier.
A pharmaceutical composition comprising a compound as claimed in claim 37 and a pharmaceutically acceptable carrier.
A pharmaceutical composition comprising a compound as claimed in claim 45 and a pharmaceutically acceptable carrier A pharmaceutical composition comprising a compound as claimed in claim 47 and a pharmaceutically acceptable carrier.
A pharmaceutical composition comprising a nucleic acid as selected from the group consisting of:
a) a nucleic acid molecule comprising the nucleic acid sequence of SEQ ID No. 7, 13, or 15, b) a nucleic acid molecule which hybridizes to a nucleic acid having a nucleic acid sequence of SEQ ID No. 7, 13, or 15, and c) a nucleic acid molecule that encodes an amino acid having the amino acid sequence of SEQ ID NO.: 8, 14, or 16.
A method for identifying proteins which interact with the proteins of claim 1, in a signal transduction pathway which promotes engulfment of apoptotic cells, comprising:
a) providing a host cell having a DNA construct comprising a reporter gene under the control of a promoter regulated by a transcription factor, which factor has a DNA binding domain and an activating domain; b) expressing in said host cell a first hybrid DNA sequence encoding a first fusion of a fragment or all of the nucleic acid of claim 2 and either said DNA binding domain or said activating domain of said transcription factor wherein the fragment regulates phagocytosis of apoptotic cells; c) expressing in said host cell at least one second hybrid DNA sequence encoding a putative interacting protein together with the DNA binding domain or activating domain of the transcription factor which is not incorporated in the first fusion; and d) determining any binding of the protein being investigated with a protein according to any of claims 1, 10 or 16 by detecting any production of the reporter gene product in said host cell.
An isolated protein from the nematode worm C. elegans which comprises an amino acid sequence of from amino acid residue 242 to amino acid residue 338 in Figure 2A or an amino acid sequence which differs from that of said protein only in conservative amino acid changes.
An isolated protein which comprises an amino acid sequence which is from amino acid 11 to amino acid 190 in Figure 20 or an amino acid sequence which differs from that of said protein only in conservative amino acid changes.
A method of diagnosis of a disorder in a patient which is associated with a defect of phagocytosis of apoptotic cells, comprising exposing a nucleic acid to a nucleic acid sample from the patient and detecting hybridization; wherein the nucleic acid is selected from the group consisting of:
a) a nucleic acid molecule comprising the nucleic acid sequence shown in SEQ ID No.: 1,3,5,7,9, 11, 13, or 15, b) a nucleic acid molecule comprising the complement of the nucleic acid sequence shown in SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15, c) a nucleic acid molecule capable of hybridizing to a nucleic acid having nucleic acid sequence according to (b) under conditions of high stringency, d) a nucleic acid molecule which encodes the amino acid which is at least 70% identical to an amino acid having the amino acid sequences of SEQ ID Nos.
2, 4, 6, 8, 10, 12, 14, or 16, e) a nucleic acid molecule which encodes an amino acid having an amino acid sequence ofSEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16.
A method of diagnosis of a disorder in a patient which is associated with a defect of phagocytosis of apoptotic cells, comprising detecting a protein in a sample from patient with an antibody to an epitope of one of the aforesaid proteins, wherein the protein is selected from the group consisting of:
a) a protein comprising an amino acid having an amino acid sequence of SEQ ID No. 8, b) a protein comprising an amino acid having an amino acid sequence of SEQ ID No. 14, c) a protein comprising an amino acid having an amino acid sequence of SEQ d) a protein comprising an amino acid having amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID Nos. 8, 14, or 16; or a sequence encoded by a nucleic acid having a nucleic acid sequence of SEQ ID Nos. 7, 13, or A protein which comprises a protein selected from the group consisting of:
a) a protein comprising an amino acid having the amino acid sequence as shown in SEQ ID No. 2, 4, 6, 8, 10, 12, 14, or 16, b) a protein comprising an amino acid having the amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID No. 2, 4, 6, 8, 10, 12, 14, or 16, c) a protein comprising an amino acid having the amino acid sequence encoded by the a nucleic acid molecule having the nucleic acid sequence as shown in SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15, wherein said protein is fused to another protein.
A protein as claimed in claim 69, wherein said other protein is an epitope tag or the product of a reporter gene.
A method for identifying whether a compound is an enhancer of a signal transduction pathway which promotes phagocytosis of apoptotic cells, comprising:
a) exposing a C. elegans, in which the expression of CED-6 is defective or otherwise suppressed, to a compound to be tested; and b) scoring for return to wild-type phenotype.
A method for determining whether a compound is an enhancer or inhibitor of a signal transduction pathway which promotes phagocytosis of apoptotic cells, comprising:
a) exposing a transgenic C. elegans as claimed in claim 17 to the compound to be tested; and b) measuring the level of phagocytic activity by scoring apoptotic corpses in the heads of L 1 larvae and/or the gonads.
An isolated protein which regulates phagocytosis of apoptotic cells, and has an amino acid sequence which is at least 70% identical to the amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16.
Use of a protein of nucleic acid for use in therapy, e.g., inflammation, autoimmune disease or cancer, comprising: a protein comprising an amino acid having the amino acid sequence of SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; a nucleic acid molecule that encodes an amino acid having the amino acid sequence of SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; an amino acid having the amino acid sequence encoded by a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15; or a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or Use of a protein or nucleic acid for use in therapy, e.g., neurodegenerative disease, stroke or sickle cell anemia, comprising: a protein comprising an amino acid having the amino acid sequence of SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; a nucleic acid molecule that encodes SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; an amino acid having the amino acid sequence encoded by a nucleic acid molecule having a nucleic acid sequence ofSEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15; or a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15.
Use of a protein or nucleic acid for the manufacture of a medicament for the treatment of a disease selected from the group consisting of: inflammation, autoimmune disease and cancer, comprising: a protein comprising an amino acid having the amino acid sequence of SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; a nucleic acid molecule that encodes SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; an amino acid having the amino acid sequence encoded by a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15; or a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or Use of a protein or nucleic acid for the manufacture of a medicament for the treatment of a disease selected from the group consisting of: neurodegenerative disease, stroke and sickle cell anaemia, comprising: a protein comprising an amino acid having the amino acid sequence ofSEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; a nucleic acid molecule that encodes SEQ ID NO.: 2, 4, 6, 8, 10, 12, 14, or 16; an amino acid having the amino acid sequence encoded by a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or 15; or a nucleic acid molecule having a nucleic acid sequence of SEQ ID NO.: 1, 3, 5, 7, 9, 11, 13, or An isolated protein which regulates phagocytosis of apoptotic cells, and has an amino acid sequence which is at least 80% identical to the amino acid sequence of 79. An isolated protein which regulates phagocytosis of apoptotic cells, and has an amino acid sequence which is at least 90% identical to the amino acid sequence of SEQ ID Nos. 2, 4, 6, 8, 10, 12, 14, or 16. 80. An isolated nucleic acid molecule that encodes a protein which regulates phagocytosis of apoptotic cells, and has an amino acid sequence which is at least 70% identical to the nucleic acid sequence of SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15. 81. An isolated nucleic acid molecule that encodes a protein which regulates phagocytosis of apoptotic cells, and has an amino acid sequence which is at least 80% identical to the nucleic acid sequence of SEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15. 82. An isolated nucleic acid molecule that encodes a protein which regulates phagocytosis of apoptotic cells, and has an amino acid sequence which is at least 90% identical to the nucleic acid sequence ofSEQ ID Nos. 1, 3, 5, 7, 9, 11, 13, or 15.
DATED this 24 th day of January 2002 Cold Spring Harbor Laboratory AND Devgen N. V.
by DAVIES COLLISON CAVE Patent Attorneys for the Applicants